Els orígens de la vegetació mediterrània
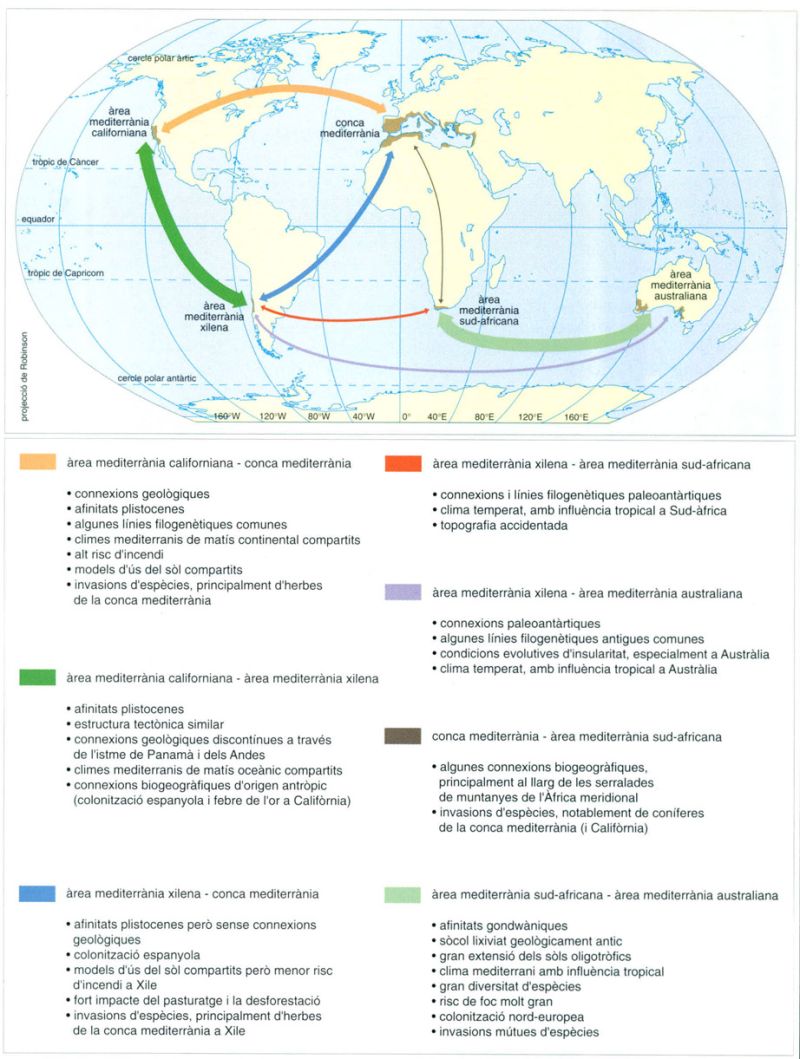
Entre les zones disjuntes de vegetació esclerofil·la de tipus mediterrani que hi ha al planeta es poden trobar diferències i similituds degudes fonamentalment a característiques geològiques i topogràfiques, a patrons de convergència evolutiva, a relacions filogenètiques i a empremtes paralleles d’intervenció humana. Les relacions filogenètiques determinen les similituds entre les dues zones de l’hemisferi occidental (Califòrnia i Xile) i les de les terres que pertanyien a l’antiga Gondwana (Sud-àfrica i Austràlia).
Editrònica
El bioclima mediterrani, tal com ja ha estat comentat, s’ha configurat en temps relativament recents. Tot i que hi ha evidències de climes amb característiques semblants a les mediterrànies durant l’última part del Miocè (7 milions d’anys enrere) i en l’època central del Pliocè (4 milions d’anys enrere), el clima pròpiament qualificable de mediterrani, amb el seu característic període d’eixut estival, va aparèixer en el Plistocè, després de la primera gran glaciació (2 milions d’anys enrere). En aquest moment, les cinc àrees que integren avui dia el bioma mediterrani ja eren clarament disjuntes i, per tant, els seus poblaments vegetals van evolucionar de manera independent els uns respecte dels altres.
En les àrees mediterrànies de l’hemisferi nord (Califòrnia i la conca mediterrània), la flora evolucionà a partir d’elements terciaris que pertanyien a ambients tropicals càlids i humits i a ambients temperats. En l’àrea xilena succeí quelcom semblant, ja que els elements terciaris preexistents pertanyien a ambients tropicals càlids i humits i a ambients antàrtics temperats i humits. En les àrees mediterrànies d’Austràlia i Sud-àfrica, la flora evolucionà exclusivament a partir d’elements tropicals, segurament comuns en totes dues àrees durant el Mesozoic. En aquestes zones, la presència de plantes que mantenen un cicle amb creixement durant l’estiu, comportament típicament tropical, reflecteix l’origen de la vegetació, tot i que les influències climàtiques tropicals que donen lloc a una certa pluviositat durant l’època eixuta han contribuït a mantenir les seves característiques.
Aquestes dues àrees mediterrànies de l’hemisferi sud comparteixen altres trets comuns, ja que els continents africà i australià, truncats cap a latituds més altes, han donat lloc a zones mediterrànies amb un grau d’aïllament superior al de les altres del bioma, cosa que s’ha traduït en una vulnerabilitat més gran davant de les invasions d’espècies forasteres. El pi de Monterey (Pinus radiata), originari de l’àrea mediterrània californiana i plantat profusament com a arbre per a fusta en totes les zones del bioma, creix molt bé en totes elles, però només s’ha comportat com a espècie invasora en les regions australianes i capenques. En la zona capenca, fins i tot algunes espècies australianes com l’arbust de la família de les proteàcies Hackea sericea, es comporten com a invasores.
L’aïllament entre les àrees i entre diferents parts d’una mateixa àrea a causa de la complexitat del relleu, sumats als repetits canvis climàtics associats a les glaciacions que han provocat moviments d’avenç i reculada de les àrees mediterrànies, han donat lloc a un panorama florístic complicat fruit d’una especiació intensa i d’un manteniment d’espècies relictuals en ambients que els són favorables.
L’elevat grau d’endemisme de la flora mediterrània es deriva d’aquests processos d’especiació en un territori fragmentat en valls i muntanyes, que es comporta com un arxipèlag en termes d’aïllament biològic.
A la regió mediterrània sud-africana, aquest procés de diferenciació florística ha anat molt més lluny que en les altres àrees del bioma, i ha donat lloc a una flora en la qual el 68% de les espècies i el 19,5% dels gèneres són endèmics. A més, la diversitat d’espècies de la flora mediterrània sud-africana és molt gran, cosa que converteix aquesta regió en la de major densitat d’espècies diferents en comparació amb d’altres àrees tropicals o temperades d’extensió similar: en una àrea de dimensions reduïdes es troben 8 500 espècies vegetals. El grau de peculiaritat de la flora de la regió mediterrània sud-africana és tan gran que constitueix per ella sola un reialme florístic, l’anomenat reialme capenc, concentrat en una regió diminuta, si el comparem amb les àrees ocupades pels altres cinc reialmes florístics considerats en el món. Així, el reialme holàrtic correspon a Euràsia i l’Amèrica del Nord, el neotropical ocupa l’Amèrica Central i gairebé tota l’Amèrica del Sud, el paleotropical la quasi totalitat del continent africà exceptuant la regió capenca, l’antàrtic, l’extrem meridional de l’Amèrica del Sud i les illes subantàrtiques i, finalment, l’australià, que correspon al continent del mateix nom.
La flora mediterrània actual evolucionà, doncs, a partir d’un conjunt d’espècies, d’ambients tropicals i temperats, que foren capaces d’adaptar-se a les noves condicions climàtiques regnants. Cal fer notar que en aquests ambients previs ja hi havia vegetals esclerofil·les que eren una resposta evolutiva a condicions d’estrès hídric o de pobresa de nutrients en els sòls. L’estrès hídric podia ser causat per factors topogràfics que provoquessin ombres de pluja o, fins i tot, inundacions durant l’època plujosa i càlida, tot obligant els vegetals d’aquests sòls a concentrar el seu creixement durant l’hivern, càlid però sec. En conjunt, aleshores, els ancestres de la flora mediterrània foren vegetals adaptats ja a condicions d’estrès hídric i nutricional que ocupaven un espai marginal en els seus respectius ambients, ja fossin tropicals o temperats. L’existència de períodes anteriors al Plistocè amb condicions climàtiques semblants a les mediterrànies amb temporada freda suau i temporada eixuta seca, encara que més suaus en l’aspecte d’ariditat, també afavorí l’aparició de vegetals especialment adaptats a la manca d’aigua.
Herbes escasses, primes i esgrogueïdes
El bioma mediterrani és dominat per comunitats vegetals llenyoses i és més aviat escàs en formacions herbàcies. Tot i que el nombre d’espècies de plantes herbàcies és considerable i representa un percentatge important de la flora, paisatgísticament les herbes tenen un paper menor. El llarg eixut estival dificulta seriosament la vida de les plantes herbàcies que, amb llurs menudes arrels, no poden arribar als nivells profunds del sòl, els únics que mantenen una certa humitat en plena època càlida. Les herbes, doncs, es veuen limitades a explotar l’aigua de les capes més superficials del sòl, totalment seques durant l’estiu. Per aquesta causa, els prats mediterranis presenten la plenitud vegetativa durant l’hivern i la primavera, mentre que a l’estiu passen un període de marcescència o repòs. Són prats secs, dominats per gramínies de fulles dures i sovint cargolades, per evitar pèrdues excessives d’aigua, que presenten quasi sempre un to clarament grogós, ben allunyat de l’aspecte verd i tendre que hom sol associar al nom de prat.
La vegetació herbàcia mediterrània
Quan la secada és extrema, les plantes herbàcies perennes tenen serioses dificultats per subsistir, i els prats desapareixen del paisatge. En aquestes condicions només poden prosperar els pradells d’anuals, herbetes minúscules, anomenades teròfits, que desenvolupen el seu cicle vital en poques setmanes durant l’època humida i que passen el període eixut en forma de llavors. Aquests pradells, sempre de reduïdes dimensions, també són els que ocupen els espais entre les herbes perennes en els prats secs o els buits deixats per petits arbustos capaços de resistir condicions de sequedat extrema. Això fa que en el bioma mediterrani la destrucció de les comunitats arbustives dels llocs més secs no doni lloc a l’aparició de prats, sinó a terrenys quasi buits ocupats només per aquests pradells de plantes efímeres, a diferència del que passa en climes més humits, on la desaparició de les comunitats llenyoses sol comportar el desenvolupament de prats substitutoris relativament esponerosos.
Les herbes mediterrànies, doncs, tenen un paper marginal en el paisatge, i ocupen llocs relativament secs i també estatges altitudinals per sobre del límit dels boscos. En determinades situacions, el foc també afavoreix la implantació de vegetació herbàcia, sobretot en els primers estadis després dels incendis, quan els arbustos que rebroten encara no han reocupat l’espai i poden aparèixer espècies herbàcies efímeres i pioneres que aprofiten aquesta disminució temporal de la competència. En llocs repetidament cremats abunden els geòfits, plantes herbàcies amb bulbs que actuen com a estructures subterrànies de resistència, que permeten una ràpida brotada i florida després del foc, com els asfòdels (Asphodelus), les orquídies (Serapias, Ophrys) i els gladiols (Gladiolus).
L’escassetat de bones pastures a l’àmbit mediterrani fa que el bestiar es vegi constret sovint a brostejar les branques baixes dels arbres o els arbustos. Òbviament, això perjudica el bon desenvolupament de les comunitats forestals, sobretot si la mancança de brots tendres empeny els pastors a calar foc a la vegetació llenyosa per tal d’afavorir l’aparició de llucs amb més palatabilitat per al bestiar; o, fins i tot, indueix a l’aparició d’herba als llocs artigats per l’incendi i subsegüentment enriquits amb nutrients per les cendres generades pel foc. Les comunitats herbàcies que, efímerament, arriben a aparèixer en aquests casos, no passen mai de prats magres i eixarreïts, fora, potser, de les primeres setmanes d’existència. De tota manera, per aquesta via la vegetació herbàcia ocupa més extensió que no li pertocaria espontàniament a l’àrea mediterrània, tant a la conca mediterrània pròpiament dita, com a les zones mediterànies de Califòrnia, Xile i Austràlia. El fenomen, altrament, ha provocat la substitució de nombroses espècies espontànies per altres de subspontànies o d’adventícies, ja que les plantes herbàcies pròpies de les tres últimes àrees estan poc adaptades a la pastura, i els pastors de les mediterrànies ultramarines, generalment d’ascendència europea, no han vacil·lat a importar espècies adequades als seus interessos pecuaris. Les formacions arbustives i arbòries obertes d’aquestes regions tenen, doncs, una flora llenyosa autòctona i una flora herbàcia en bona mesura al·lòctona, cosa que engendra paisatges de clara influència antròpica, com el “sabana oak” californià, els “espinales” xilens o les formacions de “wandoo” australianes. En l’àrea mediterrània xilena, per exemple, es pot trobar una flora amb 3 000 espècies diferents, de les quals unes 500 són introduïdes; aproximadament el 85% de les espècies introduïdes correspon a plantes herbàcies. Aquesta dominància en les espècies introduïdes també es troba en les altres zones del bioma.
Les espècies herbàcies en els àmbits mediterranis
A la conca mediterrània pròpiament dita, les plantes herbàcies perennes més abundants en el paisatge són les gramínies, com el fenàs de marge (Brachypodium phoenicoides), el llistó (B. retusum), l’espart (Stipa tenacissima) i l’espart bord (Lygeum spartum), a més de diverses espècies dels gèneres Avena, Bromus i Festuca, tot i que també hi abunden alguns elements d’origen tropical, com l’albellatge (Hyparrhenia hirta). En la zona californiana, les espècies indígenes d’Aristida, Poa i Stipa han deixat pas a les introduïdes dels gèneres Avena,Bromus i Festuca.
En la regió mediterrània xilena, en la qual predominaven les herbàcies perennes davant de les anuals, les herbàcies introduïdes també han desplaçat les natives, de manera que en algunes comunitats pradenques, un 25% de les espècies són forasteres, algunes de les quals com Erodium cicutarium, E. moschatum, Capsella bursa-pastoris, Lolium multiflorum, Stellaria media i Vulpia bromoides tenen recobriments molt importants. En l’apartat de les plantes natives cal esmentar, per la seva abundància, els geòfits, pertanyents a les famílies de les liliàcies, amaril·lidàcies i iridàcies, com també les falgueres dels gèneres Adiantum, Blechnum, Notholaena i Pellaea.
A la regió capenca són molt abundants els geòfits, com Watsonia i Gladiolus, normalment associats als llocs repetidament cremats, fins al punt que n’hi ha unes 350 espècies; també són habituals algunes espècies anuals (Dimorphoteca pluvialis, Senecio elegans), inicialment pròpies de zones degradades, que recobreixen durant la primavera amb les seves vistoses floracions. En aquesta àrea sud-africana de caràcter mediterrani, les espècies herbàcies perennes, com Themeda triandra, han estat desplaçades per herbes anuals introduïdes, bé que algunes de les autòctones, com els Oxalis, que són geòfits, tenen una presència important. Cal destacar l’extrema fragilitat i sensibilitat d’aquesta regió florística tan peculiar en la qual el problema de les plantes invasores es fa més greu, ja que mentre que en les altres àrees del bioma les espècies forasteres tenen especial èxit en les zones pertorbades, en la regió capenca les plantes introduïdes s’estenen sota totes les condicions. Així, herbes com Brachypodium distachyon, Briza maxima, Polygonum aviculare i Rumex angiocarpus només progressen en llocs alterats, i d’altres com Fumaria muralis, Lolium perenne, Spergula arvensis, Spergularia media, Silene gallica o Salsola kali ocupen tota mena d’àrees dins de la zona. A canvi d’aquesta gran susceptibilitat de la zona mediterrània sud-africana amb relació a les invasions per plantes forasteres, que han arribat a posar en perill la persistència d’algunes espècies autòctones, la regió capenca ha actuat com a exportadora de plantes herbàcies de jardí, que s’han estès arreu del món però sobretot en les altres àrees de clima mediterrani, com és el cas dels geranis (Pelargonium), dels lliris d’aigua (Zantedeschia) o de les clívies (Clivia).
En el cas d’Austràlia, les plantes herbàcies europees introduïdes, i que ara dominen el paisatge, pertanyen als gèneres Lolium i Trifolium, i a les famílies de les gramínies i de les favàcies respectivament. Les espècies originàries d’herbes pertanyen, entre d’altres, a les famílies de les poàcies, ciperàcies, asteràcies, liliàcies, orquidàcies, restionàcies i xantorreàcies. Entre les espècies natives més abundants trobem les anomenades “hummock grasses” (Triodia), molt abundants en llocs de sòls oligotròfics.
Les herbes arborescents australianes “black boys” i “black gins”
Una situació singular és la de les espècies australianes de plantes herbàcies de port arborescent. Tot i que és cert que les herbes arbòries són característiques d’Austràlia, la seva forma, de fet, no és tan estranya, ja que és similar a la d’altres monocotiledònies llenyoses arborescents, com les palmes o els Pandanus. La diferència principal és que les herbes arbòries autralianes tenen fulles molt estretes, brillants i fràgils, en comptes de les grans fulles més amples i carnoses de les palmes o pandanals. Aquesta característica esclerofil·la és una adaptació típica del medi mediterrani australià.
Les herbes arbòries, anomenades “black boys”, és a dir xicots negres (Xanthorrhoea), i “black gins”, o sigui, xicotes negres (Kingia,Dasypogon), són característiques del paisatge australià, de les zones més humides del continent, i fins i tot, bé que de manera esporàdica, del desert. Allí on es fan, resulten tan comunes que difícilment escapen a la vista, i semblaren tan estranyes als colonitzadors europeus que foren acceptades com una altra singularitat de la flora i la fauna úniques d’Austràlia, com els eucaliptus o els cangurs. La seva aparença estranya i primitiva afavorí l’aparició d’una llegenda en el sentit que serien una relíquia d’alguna antiga soca que ha sobreviscut aïlladament en el continent australià.
La configuració anatòmica i el desenvolupament
En efecte, el creixement lent de les herbes arbòries, les considerables alçades ateses i la seva aparença general primitiva, féu que hom arribés a creure que alguns exemplars d’aquestes herbes tenien fins a 6 000 anys d’antiguitat. Els darrers intents per a datar els espècimens més grossos utilitzant els moderns mètodes científics han donat uns 350-400 anys per a certs individus de Xanthorrhoea preissii i uns 600-800 anys per a uns altres de Kingia australis.
Les més típiques d’aquestes herbes arborescents tenen una mena de tronc recte, de 20-30 cm de diàmetre, i una capçada molt densa de fulles estretes. Aquestes fulles, quan envelleixen i moren, pengen i formen una faldilla per sota la capçada que persisteix fins que algun foc, usuals a la zona, l’acaba destruint. No obstant això, les bases de les fulles no es cremen fàcilment, i serveixen per a protegir del foc els brots terminals; també formen una beina externa aïlladora al voltant del tronc. Els espècimens vells poden atènyer fins a 8 m d’alçada. La major part de les espècies no presenten ramificacions, però els exemplars més vells d’algunes poden arribar a tenir-ne.
Com en qualsevol altra monocotiledònia llenyosa, les tiges de les herbes arborescents australianes presenten un cilindre central amb feixos sucosos relativament tous, envoltat per una capa més dura que consisteix principalment en grups vasculars molt atapeïts d’elements de xilema i floema, i travessats per traces de base de fulles. En la Xanthorrhoea, això és envoltat per una capa de tipus càmbium, coneguda com a dèsmium, que periòdicament origina grups vasculars secundaris internament i és protegit a l’exterior per una capa de periderma de tipus escorça. Les anelles dels grups vasculars secundaris poden agrupar-se en àrees circulars clares que alternen amb cercles foscos, la qual cosa dóna la impressió d’anelles de creixement. No cal dir que aquest procés no ocasiona el creixement en diàmetre de la tija i provoca que apareguin fissures en la beina més exterior de les bases de les fulles mortes que envolta l’epiderma. En la Kingia i Dasypogon, no hi ha dèsmium ni l’augment secundari associat a aquest, per tant la tija no pot augmentar en diàmetre, simplement s’allarga. Un aspecte curiós de la Kingia és que la seva tija produeix contínuament arrels en allargar-se, les quals surten un quants centímetres per sota del punt de creixement. Aquestes creixen cap avall a través de la zona de les bases de fulles persistent que envolten la tija.
Les formes de creixement de les flors són interessants. En les Xanthorrhoea neixen grans quantitats de floretes blanques densament atapeïdes sobre una enorme espiga terminal que surt de la capçada de l’arbre i pot créixer a gran velocitat fins a atènyer 3 m de llargada o més (s’han enregistrat percentatges d’elongació de 7 a 10 cm per dia!). El foc, bé que no és imprescindible per a desencadenar el procés, estimula la floració: l’aparença d’una herba arbòria en aquest estadi, amb un tronc negre, una capçada de fulles i una inflorescència vistosa, recordà als primers colons europeus la figura d’un caçador aborigen amb una llança, i d’aquí ve el nom de “black boy” (xicot negre). En el cas de les Kingia i les Dasypogon, les inflorescències són molt més petites i més nombroses, entre 4 i 100 per planta, situades entre les fulles a la capçada de l’arbre. Les flors individuals apareixen en caps globulars a la punta de cada inflorescència. La forma resultant recorda una dona aborigen amb elements decoratius al cap, i d’aquí el nom de “black gin” (xicota negra). Fins i tot actualment, algunes persones encara creuen que les Kingia són individus femella i les Xanthorrhoea, mascle.
L’adequació funcional i el capteniment ecològic
Tant les Xanthorrhoea com les Kingia tenen fulles dures i esclerofil·les, de secció quadrangular, amb quantitats relativament importants de feixos d’esclerènquima. Les fulles es poden trencar amb facilitat si es dobleguen manualment, però sembla que són resistents al vent. Les fulles de la Kingia són cobertes per filaments sedosos, els quals els donen un aparença argentada. Les fulles es corben cap enfora des del punt de creixement, i finalment pengen cap avall per a formar la típica faldilla. Per altra banda, en el cas de Dasypogon hookeri, les fulles no són marcadament esclerofil·les, sinó més amples —fins a 14 cm d’amplada— amb una canal còncava a l’anvers. L’examen anatòmic mostra una absència de feixos d’esclerènquima prop de la banda còncava superior de la fulla: cal fer notar que aquesta espècie creix al bosc esclerofil·le humit a la part més allunyada del sud-oest de l’oest d’Austràlia, on la característica esclerofil·la és generalment molt menys pronunciada que en altres indrets.
Les Xanthorrhoea i les Kingia presenten un esclerofil·lisme accentuat, típic de la majoria de la vegetació mediterrània australiana, que es desenvolupa a nivells més alts que en regions semblants d’altres continents. Fins i tot els deserts australians es caracteritzen per presentar pulvínuls esclerofil·les dels gèneres Triodia i Plectrachne. De manera semblant, les herbes arborescents australianes representen una forma biològica única. S’atribueix aquest fenomen a les deficiències nutritives, especialment en fosfat, molt corrents en els sòls australians. La resistència al foc és una altra adaptació que moltes plantes australianes, i en concret els “black boys”, han hagut d’adquirir.
L’origen filètic i la sistemàtica
La taxonomia de les herbes arborescents australianes encara és confusa i objecte de controvèrsia. Hom accepta l’existència de tres gèneres (Xanthorrhoea, Kingia i Dasypogon), però no hi ha unanimitat sobre a quina família pertanyen. Primer, aquestes plantes foren considerades liliàcies primitives, però d’ençà del 1829 foren englobades en una nova família, la de les xantorreàcies. Posteriorment, s’ha considerat que Dasypogon i Kingia pertanyen a una família diferent, la de les dasipogonàcies. Xanthorrhoea és un gènere endèmic australià, amb una trentena d’espècies, set de les quals creixen a la part aïllada del sud-oest d’Austràlia, una d’elles (X. thorntonii) esporàdicament al desert oriental, i la resta a la franja costanera de l’est d’Austràlia. Kingia és un monotipus amb una sola espècie (K. australis), mentre que Dasypogon comprèn tres espècies; totes del sud-oest australià.
L’hàbitat és bastant variable, i pot situar-se en el bosc humit i en el sec, en bruguerars i en aiguamolls. No tots els membres d’aquests gèneres són herbes arborescents; alguns són mates perennes, amb una ferma base subterrània que no creix cap amunt en forma de tronc. Les Xanthorrhoea arborescents són X. australis (Queensland i Tasmània), X. arborea (Nova Gal·les del Sud cap al sud de Queensland), X. johnsonii (Queensland i nord de Nova Gal·les del Sud), X. quadrangulate i X. tateana (sud d’Austràlia) i finalment X. thorntonii (part àrida de l’oest d’Austràlia). Almenys nou altres espècies produeixen troncs curts amb una alçada màxima d’1 m, i tretze més, normalment, no tenen tija. Kingia australis és arborescent, però només hi ha una espècie arborescent de Dasypogon (D. hookeri). En resum, doncs, hi ha deu espècies de plantes herbàcies arborescents en tot Austràlia.
Arbustos abundants i arbres petits
Tal com ja hem dit, el bioma mediterrani és un regne de plantes llenyoses, és a dir, d’arbres i arbustos sovint de mida modesta, però d’arrelada prou profunda per a disposar de l’aigua acumulada en les capes inferiors del sòl durant l’època eixuta. Pertanyen al tipus biològic dels faneròfits, vegetals amb gemmes situades a una alçada superior als 20-40 cm, o dels camèfits, petites mates amb gemmes situades des d’arran de terra fins als 20-40 cm. De tota manera, la distinció entre arbre i arbust no sempre és evident en l’àmbit mediterrani, car moltes espècies arbòries, ja prou baixes de mena, poden arribar a presentar-se sota una aparença netament arbustiva, tant pel que fa a la grandària, com pel que fa a la morfologia general, sota condicions ambientals limitadores. És per això que sovint trobem comunitats arbòries i arbustives amb una composició específica semblant, o que almenys comparteixen algunes de les espècies dominants. En tot cas, la riquesa florística del bioma mediterrani en arbres i arbustos és realment considerable a causa de les ja comentades peculiaritats del procés d’especiació.
Els arbres de la conca mediterrània i californians
Tant a la conca mediterrània estricta com a la zona mediterrània californiana, les espècies arbòries dominants pertanyen a les famílies de les fagàcies i de les pinàcies. En el cas de les fagàcies, la majoria de les espècies pertanyen al gènere Quercus, siguin caducifòlies com els roures, o perennifòlies com les alzines i sureres; a aquest mateix gènere, tanmateix, també pertanyen algunes espècies de port arbustiu, com el garric (Q. coccifera) de la conca mediterrània, o diverses espècies californianes del “chaparral”. En el cas de les pinàcies, es tracta majoritàriament d’espècies diverses de pins (Pinus). Tanmateix, la diversitat específica és més gran en les zones californianes que en la conca mediterrània, ja que en aquesta última les serralades transversals actuaren com a barreres en les èpoques de les glaciacions, tot augmentant les taxes d’extinció. De fet, la riquesa florística de pins, alzines i roures a la Califòrnia mediterrània és un reflex de la gran diversitat específica que aquests grups presenten a tota l’Amèrica del Nord, àrea que té una quarantena d’espècies de Quercus i una vuitantena llarga d’espècies de coníferes, trenta-quatre de les quals són Pinus.
Els pins són les coníferes més abundants a l’àmbit mediterrani estricte i al californià. Es tracta principalment de pi blanc (Pinus halepensis), pi de Calàbria (P. brutia), pi pinyer (P. pinea), pinastre (P. pinaster) i pinassa (P. nigra), a la conca mediterrània, i de P. radiata, P. sabiniana, P. attenuata i P. coulteri a la zona californiana. Altrament, cal dir que també són ben presents coníferes pertanyents a altres famílies que les pinàcies, com ara les cupressàcies o les taxàcies, per exemple. Fóra el cas de les diferents espècies de xiprers (Cupressus sempervirens al llevant de la conca, C. macrocarpa al litoral californià) i de ginebres i savines (Juniperus). A Califòrnia també destaca la presència d’alguna altra conífera, com Pseudotsuga macrocarpa, o bé de les sequoies (Sequoia sempervirens, a baixa altitud, i Sequoiadendron giganteum, a les muntanyes), i a les parts més elevades de la conca mediterrània, del teix (Taxus baccata), de cedres (Cedrus atlantica,C. libani, C. brevifolia, a l’Atlas, al Líban o a Xipre) i fins i tot d’avets (Abies pinsapo,A. maroccana, A. nebrodensis, A. cephalonica, A. cilicica, a Andalusia, al Marroc, a Sicília i Calàbria, a Grècia o a Turquia), tot i que tenen una importància paisatgística menor.
La majoria dels arbres i els arbustos planifolis californians i mediterranis estrictes pertanyen al gènere Quercus. L’absència de Quercus perennifolis a les àrees anglòfones europees ha privat l’anglès de la claredat terminològica que tenen les llengües llatines per a separar els caducifolis o roures [“robles” en castellà o “roveri” en italià], dels perennifolis o alzines [“encinas” en castellà o “querci” en italià], que són els perennifolis. En anglès, en efecte, tots acaben essent “oaks”, fet que es repeteix en francès, amb el seu únic mot “chêne” per a roures i alzines perquè l’àrea inicial d’aquesta llengua, al nord del domini occità, també està mancada de Quercus perennifolis. Per sortir del pas, hom recorre a l’artifici d’anomenar “green oak” o “chêne verte” les espècies perennifòlies, és a dir les alzines, o sigui, l’alzina pròpiament dita (Q. ilex), la carrasca (Q. rotundifolia), la surera (Q. suber), el garric i similars (Q. coccifera, Q. calliprinos) i les espècies perennifòlies californianes (Q. agrifolia, Q. chrysolepis, Q. engelmannii, Q. wislizenii i fins i tot els netament arbustius Q. palmeri i Q. morheus); les caducifòlies, és clar, no presenten cap problema, tret de ser molt escasses a l’àrea (Q. faginea, Q. pyrenaica, Q, pubescens i poca cosa més a la conca mediterrània, i Q. lobata, Q. douglasii i Q. kelloggii a Califòrnia). D’una manera general pot dir-se que els roures tenen les fulles més o menys profundament lobulades (o fins i tot incises, com és el cas de Q. kelloggii), mentre que les alzines les tenen enteres, a vegades amb els marges una mica punxents fins i tot; també són enteres les fulles del roure bru “tan oak” (Lithocarpus densiflorus), una singular alzina californiana pertanyent a un gènere de dispersió més aviat asiàtica.
Naturalment, les coníferes i les fagàcies no esgoten el paisatge arbori californià o mediterrani estricte. De la conca mediterrània fóra imperdonable ometre dos arbres que, domesticats, hi han esdevingut emblemàtics: el garrofer (Ceratonia siliqua) i, sobretot, l’olivera (Olea europaea europaea), varietat conreada de l’ullastre (Olea europaea sylvestris). I tampoc no cal oblidar els caducifolis mediterranis que, com els roures, apareixen als llocs més frescos de l’àrea o a les muntanyes, exponents dels quals foren les blades (Acer granatense, A. monspessulanum, A. macrophyllum) o el castanyer bord californià (Aesculus californica). A les riberes dels cursos d’aigua permanents de la conca mediterrània i californians també apareixen diversos caducifolis dels grups de les nogueres, els salzes i els pollancres, que seran tractats a propòsit dels sistemes aquàtics.
Els arbres mediterranis xilens

Arbres i arbustos principals del “matorral” xilè i del “chaparral” californià. La vegetació de tipus esclerofil·le comprèn la part més important de la flora de les dues àrees mediterrànies americanes. Si bé els noms que serveixen per a descriure-les suggereixen diferències entre elles —en efecte, l’àrea mediterrània californiana es caracteritza pel domini d’una espècie arbustiva del gènere Quercus, mentre que a la mediterrània xilena no hi ha cap espècie d’aquest mateix gènere— entre ambdues regions hi ha similituds, com es pot veure al dibuix. És curiós de fer notar que plantes de famílies molt diferents han adoptat estratègies similars, com és el cas de Colliguaya odorifera, de la família de les euforbiàcies, el “peumo” (Cryptocaria alba), de la família de les lauràcies, el “litre” (Lithraea caustica), una anarcardiàcia, i la quil·laia (Quillaja saponaria), una rosàcia, de la flora xilena, i la “manzanita” (Arctostaphylos glauca), de la família de les ericàcies, el “coastal live oak” (Quercus agrifolia), de la família de les fagàcies, el “chamizo” (Adenostoma fasciculatum), una rosàcia i Ceanothus leucodermis, una ramnàcia, de la flora californiana.
Eugeni Sierra
Les espècies arbòries de la regió xilena no tenen cap relació amb les que s’acaben d’esmentar. S’hi pot trobar tot un seguit d’espècies de famílies diferents com les anacardiàcies (Lithraea caustica), les rosàcies (Quillaja saponaria), les monimiàcies (Peumus boldus) o les lauràcies (Cryptocarya alba). Algunes d’aquestes espècies, com el “litre” (Lithraea caustica), que produeix irritació de pell i febre per contacte, la quil·laia (Quillaja saponaria) amb una escorça rica en saponina, de la qual es fa sabó, i el “peumo” (Cryptocarya alba) poden actuar com a dominants tant en comunitats arbòries com arbustives, tot posant de manifest una menor especialització derivada, probablement, d’un major aïllament biogeogràfic de la zona xilena. Totes aquestes espècies arbòries comparteixen amb les de la conca mediterrània i de la zona californiana un mateix aspecte d’arbre amb la capçada de color verd fosc, no gaire alt i tortuós, amb més utilitat com a llenya que no pas com a fusta, a causa de les difícils condicions de creixement.
Entre els arbres mediterranis xilens cal esmentar la presència d’una palmera endèmica (Jubaea chilensis), la més austral del món, que creix en una àrea molt reduïda de la zona mediterrània.
Els arbres caducifolis, poc abundants en la zona pròpiament mediterrània i més característics de les àrees de transició cap a climes de muntanya o més meridionals, són representats per algunes espècies de les fagàcies australs per excel·lència, pertanyents al gènere Nothofagus i que reben el nom vulgar de “robles”. La pràctica absència de coníferes és una característica compartida amb la majoria de les àrees d’Amèrica del Sud, però cal esmentar la presència del “ciprés de la cordillera” (Austrocedrus chilensis) en els vessants de la serralada andina, com també d’altres espècies que pertanyen als gèneres Araucaria, Podocarpus i Fitzroya, tot i que no són pròpiament mediterrànies sinó que corresponen a climes més freds i humits. En total, a la regió mediterrània xilena hi ha 57 espècies arbòries, de les quals 35 són endèmiques i les altres tenen una distribució més àmplia.
Els arbres mediterranis capencs
A la regió mediterrània sud-africana hi ha una única espècie arbòria autòctona (Leucadendron argenteum), pertanyent a la família de les proteàcies. Té una àrea de distribució restringida als vessants humits de la Table Mountain, prop de Ciutat del Cap, però pot considerar-se abundant, ja que ha estat profusament plantada en parcs i jardins per les seves característiques ornamentals. Altres espècies de proteàcies es troben en el límit inconcret entre l’arbre i l’arbust, però han de ser considerades més com a arbustos de gran talla que no pas com a arbres, ja que en cap cas no arriben a assolir un desenvolupament important.
En la regió mediterrània sud-africana es dóna la paradoxa que, tot i que les condicions climàtiques i edàfiques permeten el desenvolupament de masses forestals en moltes zones del bioma, la presència de vegetació arbòria indígena està molt restringida en l’espai i pot considerar-se gairebé anecdòtica. Algunes espècies arbòries forasteres, com el pi de Monterey (Pinus radiata) creixen molt bé en plantacions denses i es comporten com a invasores, tot donant lloc a masses forestals que posen de manifest la paradoxal manca d’arbres indígenes. Diverses hipòtesis expliquen aquest fet, i la més acceptada és la que fa referència als orígens de la flora sud-africana. En efecte, la possible flora arbòria de la regió capenca tindria un origen afro-montà tropical, i seria la manca en aquest “pool” inicial d’arbres pre-adaptats als sòls pobres i als incendis repetits el que donaria lloc a aquesta peculiar absència. No és gens clar per què no han aparegut espècies amb aquestes característiques, ja que el procés evolutiu podria haver donat lloc a un desenvolupament de mecanismes fisiològics d’adaptació a partir de les espècies pre-existents, però la manca total de pre-adaptacions i l’aïllament de la regió mediterrània sud-africana podrien ser possibles explicacions per a aquest fenomen.
Els arbres mediterranis australians
A la regió mediterrània australiana, la flora arbòria, com també l’arbustiva, és dominada per un gran nombre d’espècies de la família de les mimosàcies (Acacia) i de les mirtàcies, en concret del gènere Eucalyptus. De fet, tot el continent australià és dominat per espècies d’aquest gènere, capaces d’ocupar qualsevol nínxol ecològic, cosa que posa de manifest les peculiaritats biogeogràfiques d’un continent llargament aïllat.
Entre les espècies arbòries més abundants cal esmentar el “jarrah” (Eucalyptus marginata), l’arbre que ocupa la zona que es pot considerar com la mediterrània australiana típica, on fa el mateix paper que l’alzina, que l’“oak” perennifoli i que el “litre” i la quil·laia ultramarins. El “wandoo” (E. redunca) ocupa les zones de clima lleugerament més eixut, en la transició cap a les comunitats mediterrànies arbustives. A Austràlia Meridional viuen 18 espècies arbòries d’eucaliptus que formen masses forestals pures o mixtes. A la regió del sud-oest (Austràlia Occidental), però no a la del sud, apareix una zona on les pluges hivernals atenyen valors molt elevats i en la qual domina el “karri” (E. diversicolor), un arbre imposant que pot arribar a 85 m d’alçària i que forma masses forestals denses i molt desenvolupades, a vegades acompanyat d’una altra espècie de gran alçada, el “marri” (E. calophylla); juntament amb les sequoies californianes, aquests eucaliptus són els gegants d’un bioma abundant en arbres petits, fins al punt que, molt sovint, reben el qualificatiu d’arbres alguns vegetals que serien considerats arbustos en altres biomes.
Tot i que els Eucalyptus són els reis arboris en aquesta regió del bioma, cal esmentar la presència d’altres espècies d’arbres, com les pertanyents al gènere Casuarina. Les coníferes, molt rares, es restringeixen al gènere Callitris, com el “cypress pine” (C. columellaris). Finalment, cal esmentar per la seva peculiaritat les monocotiledònies arborescents pertanyents als gèneres Xanthorrhoea i Kingia, conegudes com a “black boys” i “black gins” respectivament, a causa de la forma de les inflorescències, que els donen un aspecte masculí o femení, i dels troncs renegrits pels successius incendis que han patit i als quals han sobreviscut gràcies a les adaptacions.
Els arbustos de la conca mediterrània i californians
En el cas dels arbustos, trobem una riquesa específica molt més gran en totes les àrees, però també amb diferències considerables entre elles. A la conca mediterrània hi ha un centenar llarg d’espècies d’arbustos que pertanyen a 76 gèneres i 29 famílies diferents. En termes generals, es poden diferenciar les zones oriental i occidental, dominada la primera per espècies de lamiàcies, ramnàcies i hipericàcies, i la segona per fabàcies, cistàcies i ericàcies. Alguns gèneres estan particularment ben representats entre els arbusts, i així, es troben 22 espècies de Genista, 20 de Sideritis, 12 de Cistus, Citysus, Salvia i Satureja, 10 de Thymus, 7 d’Erica i Rhamnus i 4 d’Ulex, per esmentar només uns quants exemples.
Les dimensions dels arbustos són molt variables, ja que van des d’arbrissons de 3 o 4 m d’alçada fins a subarbusts de menys de 20 cm. Paisatgísticament, són molt importants espècies com l’ullastre (Olea europaea varietat sylvestris) i el garrofer (Ceratonia siliqua), que tot i que ja hem esmentat com a arbres en estat conreat, la majoria de les vegades no ultrapassen la talla arbustiva i poden ser considerats com a tals. Altres espècies molt importants són la mata o llentiscle (Pistacia lentiscus), l’arboç (Arbutus unedo) i el garric (Quercus coccifera), per la seva dominància en comunitats arbustives altes, els brucs (Erica) i les estepes (Cistus) pel seu paper en comunitats arbustives mitjanes anomenades brolles, i les farigoles (Thymus), en el cas de les formacions arbustives de menys volada, conegudes com a timonedes. També cal esmentar la presència del margalló (Chamaerops humilis), una palmera nana que constitueix l’única espècie europea d’aquest grup.
La morfologia foliar d’aquests arbustos és tremendament variable, ja que en trobem amb fulles típicament esclerofil·les, com el garric (Quercus coccifera), amb fulles malacofil·les, com les estepes (Cistus), d’altres amb les fulles de mida molt reduïda, com els brucs (Erica), i finalment d’altres afil·les, és a dir, sense fulles i sovint amb una marcada espinescència, com és el cas d’algunes papilionàcies com ara les argelagues i gatoses, per exemple, dels gèneres Genista i Ulex.
En la regió mediterrània californiana hi ha una riquesa florística més marcada, sobretot si es té en compte que la seva extensió és clarament inferior, per les mateixes causes ja esmentades en parlar de la flora arbòria. La composició florística és ben diferent a la descrita per a la conca mediterrània, però coincideixen la majoria de les famílies, com també alguns gèneres. S’hi pot trobar una gran quantitat d’espècies arbustives que pertanyen a les fagàcies (Quercus), ericàcies (Arbutus, Arctostaphylos), rosàcies (Adenostoma, Heteromeles), anacardiàcies (Rhus) i ramnàcies (Ceanothus). La riquesa florística es tradueix en el nombre d’espècies d’alguns gèneres, com per exemple Ceanothus (25 espècies) i Arctostaphylos (19 espècies). Les espècies dominants en les comunitats típicament mediterrànies són el “chamizo” (Adenostoma fasciculatum), que té les fulles ericoides, el “scrub oak” (Quercus dumosa), amb una morfologia típicament esclerofil·la, com diferents espècies de Ceanothus i les anomenades “manzanitas” (Arctostaphylos), totes elles amb fulles també esclerofil·les. A les zones orientals de Sierra Nevada, on els hiverns són més freds i hi ha una influència desèrtica més important, és molt abundant l’arbust Artemisia tridentata. En canvi, en zones de transició cap a ambients més càlids i secs, abunden les espècies de plantes crasses de la família de les cactàcies.
Els arbustos mediterranis xilens
En el cas de la regió xilena, ja s’ha comentat la doble condició, arbòria i arbustiva, d’algunes espècies com ara el “litre” i la quil·laia. Altres arbustos abundants són Colliguaya odorifera i Kageneckia oblonga, que són típicament esclerofil·les, i Baccharis linearis, B. rosmarinifolius, Trevoa trinervis, Satureja gilliessii, Proustia cuneifolia, P. cinerea i Flourensia thurifera, que són malacofil·les caducifolis estivals. Cal esmentar també, per la seva abundància, l’“espino”(Acacia caven), un arbret amb una àrea de distribució que sobrepassa, però, l’àrea mediterrània xilena.
Un altre element distintiu d’aquesta àrea són les cactàcies columnars, per exemple el “quisco” (Trichocereus chilensis), amb exemplars que poden atènyer els 3 o 4 m d’alçària. Juntament amb aquestes cactàcies, es troben altres plantes crasses del gènere Puya, que contribueixen a donar a les comunitats arbustives xilenes un aspecte ben peculiar i diferent del de les corresponents a les altres àrees mediterrànies, excepció feta de les que es troben en les àrees més eixutes de Califòrnia, on també abunden plantes semblants.
Els arbustos mediterranis capencs
A la regió capenca, l’extrem meridional del continent africà, la riquesa florística és realment sorprenent, tal com testimonien les més de 600 espècies del gènere Erica i les més de 100 del gènere Protea. La pobresa dels sòls d’aquesta regió afavoreix l’abundància de plantes arbustives ben adaptades a aquestes condicions extremes, amb característiques morfològiques com la presència de fulles petites (Erica) o l’absència de fulles a les plantes de la família de les restionàcies, compensada per les tiges verdes que fan el paper d’òrgans fotosintetitzadors. És molt abundant l’asteràcia Elytropappus rhinocerotis, un petit arbust perennifoli d’aspecte cupressoide, al qual la sobrepastura i altres alteracions ambientals n’afavoreixen molt el seu creixement.
En els llocs més eixuts, apareixen espècies crasses afil·les o caducifòlies estivals, com per exemple Euphorbia mauritanica i Zygophyllum morgsana, o amb fulles suculentes, com Aloe ferox. Hi ha alguns arbustos esclerofil·les de talla considerable, com Olea africana, i diferents espècies dels gèneres Rhus, Euclea, Pterocelastrus i Sideroxylon, però els més abundants pertanyen a algun dels diversos gèneres de proteàcies, com Protea, Leucadendron o Leucospermum.
Els arbustos mediterranis australians
La regió australiana presenta una flora arbustiva dominada per abundants espècies del gènere Eucalyptus, tal com passa en el cas dels arbres. Hi ha una vintena d’espècies arbustives d’aquest gènere, conegudes en conjunt amb el nom de “mallee”, i amb una morfologia molt semblant, ja que tots són arbustos de gran talla o arbrets amb molts troncs que neixen d’una soca, sense que presentin un eix dominant com en el cas dels arbres. Els acompanyen moltes espècies, entre les quals altres mirtàcies (Melaleuca, Baeckea, Leptospermum) i pertanyents a les famílies de les les casuarinàcies (Casuarina), cupresàcies (Callitris), mimosàcies (Acacia), epacridàcies (Leucopogon, Astroloma), mioporàcies (Eremophila), ramnàcies (Cryptandra, Spyridium), quenopodiàcies (Atriplex, Maireana) i proteàcies (Banksia, Hakea, Grevillea), que configuren uns paisatges d’una riquesa florística considerable.
Estructura, competència i adaptacions
Les comunitats arbustives mediterrànies presenten una estructura relativament simple, feta d’un estrat arbustiu més o menys dens i un d’herbaci. Fins i tot les formacions arbustives més desenvolupades, com ara la màquia, el “matorral” o el “chaparral”, d’una densitat que les fa gairebé impenetrables, tenen una estructura molt semblant a la descrita.
L’estratificació en el cas dels alzinars
Els boscos mediterranis constitueixen un cas a part. Es presenten en les àrees més humides del bioma, i només són paisatgísticament importants en la conca mediterrània (alzinars, carrascars, suredes, rouredes), en el sud-oest d’Austràlia (“jarrah”) i en la zona californiana (“oak woodlands”), ja que són molt escadussers o totalment absents en les altres àrees. L’estructura més complexa és la dels alzinars, els boscos mediterranis més esponerosos.
L’estrat arbori pot atènyer els 20 o 25 m d’alçada, tot i que molt sovint no arriba a tant a causa de l’explotació a què són sotmesos, i perquè les fosques capçades contribueixen a crear un ambient fresc i humit a l’interior del bosc. Sota aquest sostre protector hi ha un estrat arbustiu molt ric i dens que pot assolir els 3 o 5 m d’alçada, amb plantes com el marfull (Viburnum tinus), l’aladern (Rhamnus alaternus), l’aladern fals (Phillyrea latifolia), el llentiscle (Pistacia lentiscus) i l’arboç (Arbutus unedo). Hi ha encara un estrat arbustiu baix (0,5-1 m) amb plantes de port més discret, com el galzeran (Ruscus aculeatus), l’esparreguera (Asparagus acutifolius) i la rogeta (Rubia peregrina). L’estrat herbaci, privat de llum, és molt pobre, i només duu algunes espècies com la falzia negra (Asplenium onopteris), el camedris (Teucrium chamaedrys) o la viola boscana (Viola alba).
Però la característica més singular de l’estructura dels alzinars és la presència d’un estrat lianoide ben desenvolupat, sobretot dels alzinars litorals d’alzina típica (Quercus ilex), que són els més esponerosos, no pas dels eixuts alzinars continentals de carrasca (Q. rotundifolia). Arbres i arbustos, en efecte, es veuen travats per plantes enfiladisses com l’arítjol (Smilax aspera), la vidiella (Clematis flammula), el lligabosc (Lonicera implexa), l’englantina (Rosa sempervirens) i fins l’heura (Hedera helix); en especial la vidiella, que tendeix a desenvolupar el seu fullatge penjant de les capçades dels arbres, i confereix al conjunt un sorprenent aspecte embullat. Aquests diferents estrats embolicats de lianes constitueixen, certament, boscos molt espessos, difícilment transitables, i amb un aspecte que hom podria qualificar de selvàtic, fins al punt que constitueixen el màxim nivell de complexitat vegetal dins del bioma mediterrani.
L’adequació als factors adversos
Els vegetals del bioma mediterrani han de fer front a dos problemes bàsics: la manca d’aigua durant l’època eixuta i el fred hivernal. Caldria afegir, encara que no com a problema generalitzat, la relativa pobresa de nutrients de determinats sòls, sobretot en les zones mediterrànies australianes i capenca. El fred hivernal és un factor ambiental advers que cal suportar i que pot limitar l’activitat dels vegetals, però l’aigua i els nutrients són recursos que cal obtenir del medi i que generen, per tant, relacions de competència entre espècies o entre individus de la mateixa espècie. Sequedat estival, fred i oligotròfia són tres problemes que tenen un pes i una incidència diferents segons les àrees i són, a més, tres factors d’evolució que han donat lloc a diferents estratègies per enfrontar-s’hi, ja que cadascun d’ells admet més d’una via de solució. Altrament, en un clima en què l’aigua és un bé escàs, el creixement dels vegetals es fa relativament lent i les estructures dificultosament sintetitzades són sovint protegides enfront dels herbívors, que podrien destruir en un no res allò que la planta ha tardat mesos a construir. Les adaptacions encaminades a evitar l’herbivorisme s’afegeixen, doncs, a les mencionades anteriorment.
El fred hivernal i l’activitat vegetativa
Els hiverns mediterranis no són freds sinó moderadament freds. Aquesta és una afirmació repetida constantment quan es parla del clima propi d’aquest bioma, però la veritat és que el fred crea problemes, sobretot als vegetals, incapaços com són de controlar la seva temperatura. Les espècies mediterrànies més termòfiles, per exemple el garrofer (Ceratonia siliqua) i l’ullastre (Olea europaea varietat sylvestris), tenen la seva zona de distribució molt restringida a àrees càlides, mentre que d’altres més resistents al fred com ara l’alzina (Quercus ilex) poden ocupar llocs més freds. Cal esmentar l’íntima relació que hi ha entre l’increment de la temperatura i la disminució de la pluviositat, ja que això fa que moltes espècies mediterrànies que ocupen els llocs més freds no puguin viure en els més càlids per problemes relacionats amb l’economia hídrica, i no pas amb els valors màxims de temperatura, que els són perfectament suportables.
El fred representa dos problemes diferents per als vegetals. D’una banda, en limita l’activitat, i de l’altra, pot provocar danys si les temperatures baixen per sota d’uns determinats valors. Moltes plantes mediterrànies mantenen la fotosíntesi durant una gran part de l’hivern, però no poden dur a terme activitats com la brotada, ja que la baixa temperatura actua com a limitador. La majoria de les espècies mediterrànies no poden créixer quan la temperatura és inferior a 10°C i les més termòfiles necessiten temperatures superiors als 17°C per a poder brotar. Quant als possibles danys provocats pel fred excessiu, es deriven de la formació de cristalls de glaç dins dels teixits. Això és especialment greu en el cas de vegetals perennifolis, ja que en mantenir les fulles, que són estructures poc protegides i molt hidratades, són especialment vulnerables al fred.
Molts vegetals mediterranis són especialment resistents, i l’alzina (Quercus ilex), per exemple, pot resistir temperatures de fins a -12°C sense patir danys a les fulles, i de fins a -20°C sense patir danys als troncs. D’altres són menys resistents, i així les fulles de surera (Q. suber), garric (Q. coccifera) i aladern (Rhamnus alaternus) resisteixen fins a -11°C, les de murta (Myrtus communis) i baladre (Nerium oleander) fins a -8°C, i les de garrofer (Ceratonia siliqua) fins a -7°C. La resistència al fred per una mateixa espècie o individu varia, però, al llarg de l’any i augmenta a l’hivern gràcies a una relativa deshidratació que fa més concentrats els sucs cel·lulars i vacuolars, i rebaixa el punt de congelació. En el cas de l’olivera (Olea europaea), per exemple, pot aguantar sense patir danys temperatures inferiors a -10°C a ple hivern, però només de -5°C a la tardor i la primavera. En el cas de l’alzina, aquestes diferències són de -25°C a l’hivern i de -5°C a l’estiu per als brots, i -15°C a l’hivern i -6°C a l’estiu, per a les fulles. Per contra, un caducifoli hivernal mediterrani com el roure martinenc (Q. pubescens) suporta perfectament temperatures de -35°C durant l’època en què està desproveït de fulles. Per aquestes adaptacions fisiològiques estacionals les glaçades primerenques o tardanes, rares però no absents en les regions mediterrànies, poden provocar seriosos danys en la vegetació i limitar l’àrea de distribució de determinades espècies de vida llarga encara que es presentin amb una recurrència molt baixa.
L’escassetat hídrica, la pobresa edàfica i el sistema radicular
El bioma mediterrani, caracteritzat per pluviositats moderades de distribució molt irregular, té una vegetació que combina espècies adaptades a condicions gairebé subdesèrtiques amb d’altres que necessiten condicions més humides. S’hi poden trobar tipus de vegetals tan variats com plantes crasses, caducifòlies estivals, perennifòlies i caducifòlies d’hivern. La mateixa variabilitat apareix en els sistemes radiculars, d’altra banda molt menys estudiats a causa de les dificultats metodològiques. És lògic, però, esperar detectar algunes tendències generals quant al desenvolupament i la morfologia dels sistemes d’arrels, ja que aquesta és una característica molt relacionada amb la captació d’aigua.
Les diferències morfològiques de les parts aèries es reprodueixen en les subterrànies. En àrees de “matorral” xilè instal·lades sobre sòls de més d’1 m de fondària, es detecten grans quantitats d’arrels en tot el perfil del sòl en el cas d’espècies esclerofil·les com el “litre” (Lithraea caustica), mentre que els arbustos caducifolis estivals com Satureja gilliesii limiten l’arrelada als primers 40 cm del sòl i plantes crasses com el “quisco” Trichocereus chilensis arrelen només en els primers 15 cm. Això posa de manifest la coexistència de tres estratègies diferents sota condicions ambientals anàlogues: les plantes crasses aprofiten l’aigua superficial en els moments de precipitació i l’emmagatzemen en els seus teixits; els caducifolis estivals no necessiten arrelar molt profundament, ja que la seva activitat es desenvolupa en èpoques d’humitat abundant, però exploten una capa de sòl més gruixuda ja que no són capaços d’acumular reserves d’aigua; finalment, els perennifolis, actius durant tot l’any, arrelen més profundament per tal d’assegurar-se el subministrament d’aigua al llarg de l’època eixuta.
Aquestes diferències interspecífiques tan notables responen a estratègies molt diferents, però també hi ha diferències considerables entre espècies morfològicament més semblants. En el cas dels perennifolis, per exemple, caldria suposar sempre una arrelada profunda, però en la realitat hi ha una varietat molt gran de situacions. En el mateix cas del “matorral” xilè, per exemple, Colliguaya odorifera no arrela més enllà d’1 m de fondària, mentre que el “litre” (Lithraea caustica) té arrels fins a 5 m i la quil·laia (Quillaja saponaria) pot arribar a 8 m de fondària. Semblen clars els avantatges que es deriven d’aquesta capacitat d’arrelada profunda, que ha estat detectada, directament o indirectament, en moltes espècies mediterrànies com ara Quercus calliprinos, Q. ilex, Q. dumosa, Adenostoma fasciculatum i Pistacia lentiscus entre d’altres, que poden arribar fàcilment als 8 m de fondària amb les seves arrels. L’arrelada més profunda correspon als arbres: es parla d’arrels fins a 20 m de fondària per al “valley white oak” (Q. lobata) i per al “scrub interior live oak” (Q. wislizenii) fins a 26 per al “blue oak” (Q. douglasii), i fins a 50 m per al “jarrah” (Eucalyptus marginata).
La fondària del sòl i la natura del substrat són, és clar, factors molt importants, ja que faciliten o impedeixen la penetració de les arrels, i alhora poden fer-la necessària o inútil. Els sòls poc profunds impedeixen el creixement de les arrels en fondària, però si estan instal·lats sobre un substrat calcari molt fragmentat poden fer necessària la penetració profunda d’arrels per les esquerdes. Això dóna lloc a diferències de comportament i morfologia entre individus de la mateixa espècie, segons sigui el tipus de sòl i de substrat en què han crescut. En set estudis independents sobre la profunditat d’arrelament del “chamizo” (Adenostoma fasciculatum), tres descriuen els seus sistemes radiculars com a superficials, concentrats en el primer metre del sòl, mentre que els altres quatre fan notar la capacitat de la mateixa espècie per arrelar fins a fondàries d’uns quants metres. Els sistemes radiculars són, doncs, d’una gran plasticitat perquè s’adapten a condicions edàfiques molt diferents, cosa que permet als vegetals amortir les condicions d’ariditat de determinats indrets.
Hi ha molt poques dades sobre pesos i superfícies d’arrels fines, que són les realment funcionals en la captació d’aigua i nutrients. La relació entre superfície d’arrels funcionals (superfície absorbent) i la superfície de fulles (superfície transpirant) pot ser un molt bon indicador de l’estratègia de les diferents espècies, però el nivell actual de coneixements no permet fer generalitzacions. Se sap que només el 10% en pes de les arrels correspon a arrels fines i, d’aquesta fracció, només una part actua com a plenament funcional. En general, la superfície de sòl ocupada pel sistema radicular és superior a la de capçada i varia segons la fondària del sòl. Per al “chamizo” (Adenostoma fasciculatum), per exemple, s’han trobat relacions de superfície d’arrelada/superfície de capçada de 2 o 3 per a sòls profunds, i de 7 per a sòls prims; això posa de manifest la plasticitat ja esmentada en parlar de la fondària d’arrelament. En tot cas, tot i la manca d’informació sobre aquest aspecte, cal remarcar la presència relativament abundant d’adaptacions dels sistemes radiculars que afavoreixen una absorció de nutrients molt efectiva, com per exemple l’abundància d’arrels fines en la capa més superficial del sòl i la virosta, la micorització o la simbiosi amb bacteris fixadors de nitrogen. Aquestes adaptacions, presents en tots els biomes però especialment abundants en condicions d’oligotròfia edàfica, responen a la relativa abundància dels sòls pobres en el bioma mediterrani.
La relació entre la biomassa de les arrels i la de les parts aèries també és un bon indicador de l’estratègia de cada espècie. Les diferents espècies mediterrànies estudiades donen valors molt variables, ja que poden estar molt influïts per la repetida destrucció de les parts aèries per incendis o tales. En general, s’esmenten valors per a aquesta relació entre 0,2 i 0,9 per a arbres i entre 1 i 6 per a arbustos. Les excepcions són, però, molt abundants, i així, en alzinars trobem valors que oscil·len entre 0,4 i 1,2 segons que dominin els arbres de gla o els de rebrot. En el cas d’arbres, s’han trobat valors puntuals molt més elevats, per exemple de 2,3 en el “peumo” (Cryptocarya alba) i de 5 en el “litre” (Lithraea caustica). Cal fer notar que aquests valors tan elevats són, segurament, producte de l’eliminació repetida de les parts aèries i que no són exclusius del bioma mediterrani. En boscos temperats americans, per exemple, es donen valors superiors a 6 per a arbustos caducifolis com el “scrub oak” (Quercus ilicifolia), en períodes de regeneració després d’incendis. En tot cas, la combinació de la secada estival amb la presència periòdica del foc, fa esperar que els elevats valors de biomassa subterrània siguin molt freqüents, encara que no exclusius, en els biomes mediterranis. La coexistència d’estratègies diferents quant a l’arrelada permet un aprofitament més exhaustiu dels recursos hídrics i nutricionals dels sòls i alhora contribueix a disminuir les relacions de competència, ja que diferents espècies exploten capes distintes del sistema edàfic.
Molts arbres i arbustos mediterranis amb capacitat per a rebrotar presenten unes estructures semisubterrànies en forma de soques engruixides i formades per un teixit anomenat lignotúber, capaç d’emmagatzemar aigua i nutrients i de donar lloc a nous brots quan les parts aèries han estat eliminades per foc, pastura o tala. Aquestes estructures, doncs, estan relacionades tant amb el foc com amb l’economia hídrica i de nutrients, i tenen un paper molt important en l’ecologia i el funcionament dels sistemes mediterranis.
L’eixut estival i el sistema foliar
Els vegetals mediterranis han de suportar eixuts prolongats precisament durant l’època càlida, cosa que determina tot un seguit d’adaptacions englobades sota el nom de xerofitisme. Davant d’un problema com aquest, i segons la durada del període eixut, hi ha dues estratègies bàsiques: mantenir les fulles tot reduint les pèrdues d’aigua per control estomàtic, o perdre les fulles durant el període sec tot limitant l’activitat a l’època més fresca i humida. L’adopció d’una o altra estratègia és funció de l’economia dels recursos, i així, quan els costos de manteniment d’una fulla, en termes de quantitats de carboni necessàries, durant el període eixut són menors que els costos de fabricació d’una fulla nova, la perennifolietat és la millor estratègia. Evidentment, els costos de manteniment i fabricació d’una fulla depenen de la seva estructura i no són quantitats fixes; però, en termes generals, es pot dir que quan la durada del període eixut és superior a 100 dies s’afavoreix l’estratègia dels caducifolis estivals, i quan és inferior, la dels perennifolis. En condicions intermèdies, conviuen totes dues estratègies, però en condicions extremes domina l’una o l’altra.
A més de sobreviure a la secada, una fulla ha de durar un temps suficient per a recuperar el seu cost de construcció, el qual depèn de l’estructura i la composició bioquímica. Això fa que ens trobem amb longevitats foliars molt variables, ja que molts caducifolis estivals recuperen el cost de construcció foliar en només 90 dies, mentre que molts perennifolis necessiten 1,2 o fins i tot 3 anys per a recuperar-lo. Com que la taxa de fotosíntesi varia segons la quantitat de llum rebuda, trobem temps de recuperació dels costos de construcció i longevitats diferents, fins i tot per a una mateixa espècie i individu, segons la posició de la fulla en la capçada de l’arbre o de la situació de l’individu en un solell o una obaga. Així, per exemple, la longevitat foliar del “chamizo” (Adenostoma fasciculatum) és de 2 a 3 anys en les obagues i de només d’1 a 1,5 anys en els solells.
La longevitat foliar també està relacionada amb la disponibilitat de nutrients essencials com el nitrogen, ja que l’oligotròfia dels sòls afavoreix l’estratègia perennifòlia. En condicions de manca de nutrients és millor construir fulles més duradores, és a dir, estructuralment més resistents i menys atractives per als herbívors. És interessant fer notar que la secada estival s’afronta amb fulles d’una longevitat propera a un any, mentre que la manca de nutrients pot estimular longevitats superiors a un any. A més, les fulles producte d’una oligotròfia tenen un alt percentatge de materials estructurals, tal com passa en les que han evolucionat esperonades per la secada, ja que han d’evitar les deformacions i els danys mecànics que la pèrdua d’aigua pot produir. Finalment, i per complicar més el panorama d’interaccions, l’elevada relació C/N de les fulles riques de materials estructurals les fa especialment resistents a la descomposició, cosa que fa que una vegetació perennifòlia resultat d’una adaptació a condicions de secada estival pugui donar lloc a una relativa pobresa de nutrients a causa de la baixa velocitat de descomposició del material vegetal foliar.
Això explica que part de la flora mediterrània hagi pogut evolucionar a partir d’elements anteriors adaptats, no a la secada estival, sinó a la manca de nutrients, factor que pot donar-se sota qualsevol clima. També confirma aquest conjunt de consideracions el fet que la majoria dels vegetals mediterranis xilens, que creixen sobre sòls relativament rics, tenen una taxa de renovació foliar alta, és a dir, una longevitat foliar determinada climàticament i relativament baixa. En la regió californiana, amb sòls més pobres i condicions climàtiques de secada més extremes, la longevitat foliar té tendència a ser més gran. Finalment, a Sud-àfrica i Austràlia, els efectes climàtics extrems es veuen reforçats per una marcada oligotròfia dels sòls, tot donant valors de longevitat foliar encara més gran.
La morfologia foliar també és molt important com a factor d’adaptació a les condicions de secada estival. Els caducifolis estivals tenen fulles malacofil·les, és a dir, relativament amples i poc dures, ja que, de fet, eviten el problema de l’eixut estival passant-lo en estat de latència i inactivitat. Molts presenten fulles cobertes de pilositat que evita les pèrdues excessives d’aigua: com que no són caducifolis estrictes, han de protegir-se de la dessecació. En efecte, mentre que alguns perden totes les fulles, d’altres es limiten a reduir la superfície foliar i l’activitat segons la manca d’aigua, exhibint un comportament relativament plàstic. Aquest seria el cas d’arbustos com ara les estepes (Cistus) o l’arçot (Rhamnus lycioides), que tot i reduir la seva àrea foliar considerablement i poder fins i tot presentar fulles relativament més petites i dures a l’estiu, mai no arriben a quedar totalment despullats.
Els perennifolis, en canvi, han d’afrontar el problema directament i fan servir estratègies variades. Els esclerofil·les tenen fulles fortament esclerificades, riques de materials estructurals com la lignina i la cel·lulosa i sovint recobertes d’una cutícula dura. Són fulles endurides, cosa que les fa estructuralment resistents a la dessecació, i amb una alta capacitat de regulació estomàtica que els permet deixar de transpirar quan disminueix l’arribada d’aigua. La quantitat d’aigua que contenen les fulles, menor que en el cas dels malacofil·les, també és més constant a causa de la seva capacitat de regulació. Els vegetals esclerofil·les, molt difosos en el bioma mediterrani, poden presentar fulles relativament grans; és a dir, són planifolis, com molts Quercus, proteàcies o la majoria dels arbres i els arbustos xilens. D’altres presenten fulles molt petites i recobertes de pilositat anomenades fulles ericoides, tal com succeeix en les ericàcies i epacridàcies, per esmentar-ne alguns exemple. Són considerades també esclerofil·les les gramínies perennes com ara l’espart bord (Lygeum spartum) o els pelaguers (Stipa), que tenen fulles llargues i recargolades, de secció gairebé cilíndrica, per a evitar la dessecació.
Els xeròfits afil·les són els que han perdut les fulles i tenen tiges verdes fotosintetitzadores, cosa que els permet de reduir les pèrdues d’aigua perquè presenten una superfície de transpiració molt petita. Aquest seria el cas de les restionàcies sud-africanes i d’alguns arbustos de la conca mediterrània com la ginesta (Spartium junceum). L’estratègia més dràstica, poc corrent en el bioma mediterrani, consisteix a perdre totalment les fulles i desenvolupar teixits per emmagatzemar l’aigua que cau durant el període humit, o bé convertir les mateixes fulles en magatzems d’aigua. Aquestes, les anomenades plantes crasses, són típiques de climes més eixuts i irregulars que el clima mediterrani, com ara els climes subdesèrtics, però en trobem alguns exemples en el bioma, que van des de petites plantes herbàcies com els crespinells (Sedum) fins els grans cactus columnars xilens (Trichocereus chilensis), passant per arbustos de mida moderada, com les lletereses sud-africanes (Euphorbia) o les “spanish bayonet” (Puyucca) californianes, de fulles crasses i punxents com les baionetes espanyoles que els han donat nom.
La defensa contra l’herbivorisme
Una fulla de longevitat considerable, costosament fabricada en un medi on l’aigua és un bé escàs, és quelcom preciós que cal protegir dels herbívors, que poden destruir en pocs dies el producte de l’activitat de mesos. L’espinescència es relaciona amb la defensa contra els herbívors. L’acumulació d’essències volàtils en les fulles i les tiges contribueix d’una manera efectiva a fer que el material vegetal sigui poc apte per a ser consumit per herbívors, tot i que alguns animals típicament mediterranis, com les cabres, són capaços de menjar-s’ho gairebé tot, per aromàtic i amargant que sigui. Moltes espècies mediterrànies de lamiàcies (Thymus,Salvia, Rosmarinus, Lavandula), de cistàcies (Cistus,Helianthemum), de pinàcies (Pinus), de cupressàcies (Cupressus) i de mirtàcies (Myrtus, Eucalyptus), per esmentar només les més oloroses, estan carregades d’essències que confereixen als paisatges mediterranis aquesta flaire intensa i característica, una aroma que esdevé realment forta durant els mesos secs i calorosos, i que no prové de les floracions sinó de les essències acumulades en les fulles i les tiges.
L’acumulació d’essències també pot tenir altres interpretacions relacionades amb l’economia hídrica, amb el pirofitisme o amb la competència. En evaporar-se, les essències augmenten la densitat de l’aire dins les cavitats de la fulla i sobre ella mateixa, i fan que augmenti la resistència a la difusió del vapor d’aigua des de les parets cel·lulars i disminueixi així la transpiració. A més, com que fan que augmenti la inflamabilitat de la planta, poden interpretar-se com una més de les característiques que configuren la síndrome adaptativa del pirofitisme.
Finalment, en alguns casos s’ha comprovat que les substàncies aromàtiques acumulades en els òrgans vegetals passen al sòl quan les plantes es descomponen i llavors actuen inhibint la germinació d’altres espècies, tot evitant la competència per l’aigua i l’espai. Aquest fenomen, típic de zones àrides, s’ha comprovat en els eucaliptus (Eucalyptus) i en algunes espècies del “chaparral” com Salvia leucophylla i d’altres que pertanyen als gèneres Arctostaphylos i Adenostoma. Aquests inhibidors de la germinació tenen una vida que va de menys d’un any fins a diversos anys, segons les espècies que els produeixen, i això assegura l’eliminació de la competència fins i tot en el cas que les parts aèries de la vegetació siguin eliminades per alguna causa.
El foc i la vegetació
El foc és un factor ecològic de gran importància en diferents biomes, no només en el mediterrani. Els boscos boreals de coníferes, per exemple, pateixen incendis cíclics amb una periodicitat propera als 200 anys des de fa mil·lennis, tal com ho testimonien les capes de cendra acumulades en els sediments lacustres. Les praderies també pateixen incendis amb una periodicitat menor, cosa que permet explicar la dominància d’una vegetació herbàcia sota condicions climàtiques favorables per al desenvolupament de masses forestals. Però en cap altre bioma el foc no té tanta rellevància ecològica com al mediterrani.
Les causes de l’incendi espontani
En el bioma mediterrani el foc ha tingut i té una gran importància com a factor ecològic, ja que la coincidència de l’època càlida amb l’eixuta representa una combinació que afavoreix els incendis. El mateix caràcter de la vegetació esclerofil·la fa que sigui especialment inflamable, i per això facilita encara més l’aparició i l’extensió dels incendis. A més, els focs rarament cremen estructures vegetals de diàmetre superior als 2 cm, cosa que provoca una acumulació de biomassa morta i semicarbonitzada que representa un augment de la inflamabilitat mentre no desaparegui per descomposició. La causa natural més important dels incendis són els llamps, molt corrents a les zones mediterrànies, sobretot durant les curtes i violentes tempestes estivals formades per inestabilitzacions locals de masses d’aire; aquestes tempestes, molt riques en aparell elèctric, poden deixar quantitats minses o nul·les de precipitació, de manera que encara fan més factible l’inici de l’incendi. Cal esmentar com a excepció la zona mediterrània xilena, en la qual són molt rares les tempestes amb aparell elèctric. En la regió californiana, en canvi, els llamps són un factor d’incendi de primera magnitud i s’hi han arribat a enregistrar fins a 1 750 incendis (l’any 1972) en les àrees ocupades pels boscos.
La freqüència d’incendis en condicions de baixa o nul·la humanització és difícil d’avaluar, però s’han fet estimacions aproximatives que parlen de focs amb recurrències de 15 a100 anys per al “mallee” australià, de 6 a 40 anys per al “fynbos” capenc i de 25 a 50 anys en el “chaparral” californià, tot i que en aquest últim cas les formacions arbustives més seques i veïnes de prats secs poden patir incendis amb una periodicitat anual. En el cas de la regió mediterrània xilena, hi ha poca informació sobre la freqüència d’incendis anterior a la humanització, però la raresa de les tempestes amb aparell elèctric fa pensar en una recurrència baixa. Alguns autors estimen la recurrència dels incendis en l’àrea xilena de prop de 270 anys, però sorprenentment, en aquesta regió abunden les espècies amb capacitat de rebrotar després del foc, cosa que s’interpreta com una adaptació a aquest factor i que fa pensar que, efectivament, té una relativa importància. En la conca mediterrània, la intensa i antiquíssima humanització combinada amb la complexitat estructural del paisatge, fan gairebé impossible estimar la freqüència dels focs naturals, a no ser que solucionem el problema tot considerant que els humans són el factor natural més important quant a l’aparició dels incendis ja des d’èpoques molt antigues.
Un factor ambiental d’efectes tan dràstics i recurrents com el del foc ha deixat la seva marca en la vegetació mediterrània. Cal no menysprear els efectes dels incendis en les èpoques en què l’espècie humana no tenia el paper preponderant dels últims mil·lennis, ja que tot i que els focs devien ser menys freqüents, les àrees afectades podien ser molt més grans, ja que el foc actuava sobre un paisatge molt més homogeni que l’actual, solcat de barreres (conreus, carreteres) que actuen com a tallafocs. Paradoxalment, tal com ja tornarem a comentar, en algunes àrees mediterrànies hi ha actualment un gran nombre d’incendis, però la freqüència amb què cada àrea és cremada pot ser menor que en temps antics.
Les adaptacions al foc: rebrotar o germinar a l’engròs
A conseqüència de la recurrència dels incendis, en el bioma mediterrani trobem moltes espècies vegetals que presenten adaptacions encaminades a resistir el foc, tot i que algunes han anat més enllà i han desenvolupat mecanismes reproductius i cicles vitals que donen clars avantatges a l’espècie quan el foc fa acte de presència. Els vegetals del primers tipus reben el nom de piroresistents, mentre que les segones són anomenades piròfits. Els casos més extrems de pirofitisme suposen un conjunt d’adaptacions que fan a la planta especialment inflamable, és a dir, que afavoreixen l’aparició i l’extensió d’incendis.
La regeneració per rebrotada: piroresistència
Un dels principals mecanismes de piroresistència és el desenvolupament d’escorces gruixudes i poc inflamables, que actuen com a aïllants tèrmics tot protegint meristemes subcorticals capaços de rebrotar i donar lloc a noves fulles. Algunes espècies d’eucaliptus, com el “jarrah” (E. marginata) i el “marri” (E. calophylla), i també diverses de Quercus presenten aquesta adaptació; el cas més accentuat és el de l’alzina surera (Q. suber), un arbre típic de la part occidental de la conca mediterrània que presenta una escorça suberificada i molt poc inflamable, d’uns quants centímetres de gruix, que permet la rebrotada de branques amb diàmetres més grans de 2 o 3 cm, tot donant lloc a una ràpida regeneració de les capçades després dels incendis. La temperatura assolida durant l’incendi i el gruix de l’escorça determinen el diàmetre de les branques que han tingut els teixits subcorticals efectivament protegits i que, per tant, podran rebrotar.
L’escorça és un aïllant tèrmic eficaç, i el seu gruix és el factor més important per a determinar la resistència al foc. A la conca mediterrània la surera (Quercus suber) produeix grans quantitats de rebrots a les branques quan l’incendi no ha estat gaire intens, o quan no se li ha llevat l’escorça (el suro), i reconstitueix, en un termini de 2 a 3 anys, la massa foliar de l’arbre. Nombroses espècies d’eucaliptus (Eucalyptus) posseeixen escorces resistents al foc, les quals els permeten una ràpida recuperació del fullatge després dels incendis. Aquests arbres es caracteritzen per una escorça generalment gruixuda, que protegeix el càmbium i dóna resistència al tronc. Els brots proveïts de fulles provenen directament dels borrons en repòs que hi ha sota l’escorça del tronc o de les branques. Certs pins, com ara el pi de Canàries (Pinus canariensis), també presenten excepcionalment aquesta capacitat de sobreviure.
Molts arbustos mediterranis presenten bases de troncs engruixides i semienterrades amb capacitat per a rebrotar. El 50% de les espècies dominants del “chaparral” californià, el 65% de les del “fynbos” capenc, el 70% de les que constitueixen els boscos australians de “jarrah” i la gran majoria de les llenyoses de la conca mediterrània, coníferes excloses, són capaces de rebrotar. En aquests casos les parts aèries es cremen totalment però es manté viva la soca o rabassa, gràcies a l’aïllament tèrmic que comporta el sòl que la recobreix parcialment. Pensem que en el cas d’un incendi que assoleix temperatures aèries de 800 a 1 100°C, a 5 cm de fondària la temperatura del sòl no puja de 40°C i, a 10 cm de fondària, pràcticament no canvia. Aquestes soques, constituïdes per un teixit especialitzat anomenat lignotúber, també actuen com a magatzems d’aigua i nutrients i asseguren la rebrotada ràpida fins i tot en èpoques estivals eixutes, sense necessitat d’esperar les pluges tardorals o hivernals. Molts vegetals que presenten aquesta estructura, la desenvolupen ràpidament en els primers anys de la seva vida, durant els quals destinen gairebé tots els recursos al desenvolupament de la soca i les arrels. Els “jarrah” (Eucaliyptus marginata), per exemple, ja tenen el lignotúber ben desenvolupat cinc anys després d’haver germinat, cosa que n’assegura la supervivència en el cas que hi hagi foc.
La major part dels arbres de les bosquines, garrigues i màquies europees i els del “chaparral” californià produeixen rebrots que neixen de les soques. Cada espècie disposa d’un ventall de possibilitats de resistència al foc que varia segons la intensitat d’aquest, possibilitats que poden variar, al seu torn, segons els estadis fenològics al llarg de l’any i/o l’estadi de creixement de la planta. Així, l’alzina (Quercus ilex), que rebrota abundantment de soca, pot emetre rebrots per mitjà de borrons subjacents a l’escorça quan l’incendi és poc intens, i els individus presenten una escorça prou gruixuda. Amb l’excepció, potser, d’una quinzena d’espècies, tots els Eucalyptus tenen lignotúbers; aquestes excrecències llenyoses s’inicien amb petites creixences a nivell de les axil·les dels cotilèdons i de les primeres fulles de les plàntules. A mesura que la planta creix, el lignotúber també ho fa i s’enterra progressivament dins el sòl. En els Eucalyptus arbustius, el lignotúber pot créixer contínuament. Aquest òrgan és una font de borrons vius que poden sobreviure després d’un incendi gràcies a la seva posició subterrània. Així mateix, hi ha nombrosos brucs (Erica) que tenen un tipus de rabassa similar, la qual els permet de rebrotar abundantment i reconstituir amb rapidesa la part aèria destruïda pel foc.
En determinats casos no es tracta de rabasses semienterrades, sinó de tiges subterrànies totalment protegides pel sòl. Aquesta adaptació es presenta en alguns arbustos, com el garric (Quercus coccifera), i també en plantes herbàcies dotades de bulbs, com per exemple els Gladiolus, Asphodelus i Ophrys. Els vegetals d’aquest darrer grup, els anomenats geòfits, són extremament abundants en comunitats repetidament afectades pel foc; alguns, només floreixen després dels incendis, mentre que si no hi ha foc es limiten a tenir creixement vegetatiu. En el cas de plantes herbàcies com les gramínies, les bases foliars, molt rígides, actuen com a protectores i permeten a la planta rebrotar i regenerar-se ràpidament.
Un cas singular és el dels “black boys” o “yacca” australians (Xanthorrhoea), plantes herbàcies de port arbori ben adaptades a la rebrotada després dels incendis. Les arrels dels plançons d’aquestes herbes arborescents són contràctils, i dirigeixen les delicades zones de creixement terra avall. En el decurs d’un foc, una planta jove perd les fulles, però es renova des de baix. La tija d’un plançó jove s’afiança sota terra i poden passar molts anys abans que una tija aèria no sobresurti del sòl. Primerament cal que la base de la tija arribi a un diàmetre adult complet abans que es produeixi l’elongació. Quan això ocorre, el punt de creixement és protegit de l’escalfor de qualsevol foc per les bases de les fulles, que són atapeïdes i, per tant, menys inflamables que la capçada de fulles que es troben exposades a l’aire lliure. Les bases de les fulles són resinoses i, per tant, inherentment inflamables, però com que la majoria dels incendis d’arbustos són transitoris, normalment resisteixen al foc durant temps suficient.
El creixement dels llucs en qualsevol espècie rebrotadora és sempre molt ràpid, ja que disposen d’un sistema radicular ben desenvolupat que els permet captar aigua i nutrients, i poden atènyer alçades de fins a 1 m en el període en què una plàntula nascuda de llavor no passaria d’uns pocs centímetres. Aquest creixement sol ser molt actiu durant els primers 4 o 5 anys, després s’alenteix i esdevé gairebé nul uns 25 o 30 anys després de l’incendi. La morfologia dels rebrots és variada i condiciona el recobriment del sòl, o sigui, la seva protecció davant de l’erosió. La majoria dels rebrotadors de soca donen lloc a tanys que creixen verticalment i agrupats, de manera que recobreixen poca superfície. Els que rebroten de soca i arrels o bé a partir d’estolons subterranis donen lloc a un recobriment molt important, tot i que no solen atènyer les alçades dels anteriors. En aquest últim cas, tal com succeeix en les garrigues de garric (Quercus coccifera), la regeneració és rapidíssima i l’autosuccessió assegura la pervivència de la comunitat malgrat la repetida acció del foc.
La possibilitat d’emetre rebrots podria ser una adaptació antiga, que només aparegués quan el fullatge fos destruït per un agent extern. Mentre el fullatge es mantingués viu, hi hauria una inhibició que privaria que els borrons entressin en activitat; però en desaparèixer la part aèria, aquesta inhibició també desapareixeria, i els borrons en repòs produïrien nous rebrots. El pas del foc pot estimular les plantes resistents perquè produeixin inflorescències, però aquest és un fenomen excepcional en els vegetals de la conca mediterrània, bé que ha estat observat en les monocotiledònies i estudiat concretament en Xanthorrhoea australis.
La regeneració per germinació: antracofitisme i pirofitisme
Alguns vegetals moren en ser cremades les seves parts aèries. Aquest és el cas d’alguns petits arbustos, de moltes plantes herbàcies i d’algunes arbòries com els pins. En aquest cas, els individus morts són substituïts per nous individus nascuts de llavor, que troben un espai sense competència i adobat per les cendres. Aquesta estratègia afavoreix les espècies amb una gran capacitat de producció de llavors i amb una també gran capacitat de dispersió. Així mateix, les espècies més afavorides són les de caràcter heliòfil, ja que l’elevada insolació que es deriva de la destrucció de la coberta vegetal n’afavoreix la germinació. La destrucció per combustió de les substàncies inhibidores de la germinació que es desprenen dels òrgans vegetals de determinades espècies durant el procés de descomposició pot afavorir també l’aparició de nous individus nascuts de llavor.
Alguns autors distingeixen dues estratègies quant a la regeneració per llavors per tal de diferenciar clarament el veritable pirofitisme. Els antracòfits són, senzillament, espècies oportunistes amb elevada capacitat de dispersió que colonitzen ràpidament els llocs cremats a partir de llavors produïdes en zones no afectades pel foc, tal com fa per exemple la calcida blanca (Galactites tomentosa), que produeix una gran quantitat de fruits amb dispersió anemòcora. En canvi, els vegetals que es regeneren per llavor i que, a més, la seva germinació és afavorida pel foc, són els anomenats piròfits. Presenten característiques com una producció elevada de llavors fortament protegides per cobertes dures i que germinen amb dificultat si no ha sofert un xoc tèrmic, i acumulació de substàncies volàtils que les fan especialment inflamables, tot i que aquesta característica pot ser interpretada com una adaptació relacionada amb l’economia hídrica.
Constitueixen un bon exemple d’aquesta estratègia les estepes (Cistus), arbustos heliòfils que produeixen una gran quantitat de petites llavors protegides per una coberta dura i que germinen massivament després de l’incendi: en un any sense foc la germinació de les estepes pot ser de prop de 10 o 20 individus per metre quadrat, mentre que després d’un incendi pot assolir valors de 300 o 400 individus per metre quadrat. Les plantes adultes moren, i és per això que la possibilitat de recolonització depèn de la freqüència d’incendis, que ha de permetre el desenvolupament i la floració dels nous individus.
La majoria d’arbustos piròfits, però, poden produir llavors a l’any d’haver germinat, cosa que n’explica la dominància en els paisatges repetidament cremats i la progressiva desaparició, si el foc no fa acte de presència. Per exemple, l’espècie californiana Lotus scoparius es comporta com un piròfit germinador: es fa molt abundant fins a 5 anys després del foc i queda reduïda a uns pocs individus esparsos al cap de 20 anys, tot i que les llavors acumulades en el sòl n’asseguren la nova preponderància, si torna a fer aparició el seu aliat. Són, doncs, vegetals que afavoreixen l’aparició i l’extensió del foc i que colonitzen ràpidament les zones cremades gràcies a una abundant germinació. L’únic problema seriós que tenen és el derivat de freqüències d’incendi massa altes, ja que en aquestes circumstàncies no tenen temps de desenvolupar el seu cicle complet entre dues pertorbacions i no poden, per tant, produir noves llavors. En els casos més exagerats de pirofitisme, les llavors romanen sobre la planta i només són alliberades per efecte del foc, protegides per estructures que les aïllen de les altes temperatures: és el cas de les plantes piròfiles, tractat més endavant.
La germinació experimental mostra que la resposta a l’escalfor depèn, alhora, de la temperatura a què són exposades les llavors i de la durada de l’exposició. Períodes d’exposició llargs a temperatures relativament baixes poden donar el mateix resultat que exposicions curtes a temperatures altes, i la resposta tant pot ser la germinació com la mort de la llavor. El resultat, però, també depèn de l’espècie de les llavors i, per tant, dels seus caràcters físics i fisiològics. Així, les llavors de l’estepa negra (Cistus monspeliensis) presenten l’índex de germinació més alt quan han estat sotmeses a temperatures de 90 a 150°C; no obstant això, calen 15 minuts d’escalfament a 90°C per assolir aquests índexs de germinació, mentre que a 150°C només cal un minut. A partir d’aquests valors, les temperatures són massa altes i maten l’embrió. Aquests resultats han estat observats en espècies esclerofil·les australianes, en espècies del “chaparral” californià i en plantes de la conca mediterrània.
La major part d’espècies de pins (Pinus), incapaces de rebrotar, només poden refer-se dels incendis germinant a partir de llavors. Tota la seva estratègia s’adequa, per tant, a explotar aquesta habilitat. Les llavors, per exemple, del pi blanc (P. halepensis), arbre que forma grans boscos a les regions central i occidental de la conca mediterrània, són disseminades mecànicament en esclatar amb força les pinyes durant o després del pas del foc. Per això, després de l’incendi, les pinedes són colonitzades per una gran quantitat de plàntules de pi, que reconstitueixen una nova pineda. Atesa l’aptitud d’aquestes llavors per a germinar sota el sòl mineral de les zones totalment exposades a la llum, on la competència és més baixa i els elements minerals són abundants, les pinedes poden perpetuar-se així indefinidament, sempre que la freqüència dels incendis no sigui gaire elevada, i doni als arbres prou temps per a produir pinyes fèrtils.
En la natura, la resposta de l’estoc de llavors colgades al sòl depèn no solament de les propietats de les llavors, sinó de les característiques del foc, és a dir de la seva intensitat, de la calor que penetri dins el sòl i del temps d’exposició, i també de la profunditat a què es trobin les llavors. Després d’un incendi, nombroses espècies herbàcies, anuals o bianuals, apareixen amb gran abundància, floreixen i desapareixen en el termini de 2 o 3 anys. Aquest comportament ha estat observat en moltes regions: a Califòrnia, a Austràlia, a l’Àfrica capenca, a Israel, a França, etc. En el “chaparral” californià, hom ha estudiat a fons aquest tipus de fenomen i s’ha arribat a la conclusió que la profusió de germinacions i el creixement d’espècies herbàcies és causat per la suspensió d’una inhibició química, induïda per l’arbrat, que actua sobre les llavors colgades al sòl. En aquest cas, no són limitacions de llum, d’aigua o de nutrients, doncs, el que obstaculitza la germinació, sinó fenòmens d’al·lelopatia. Extractes procedents de les espècies llenyoses afecten la germinació de les llavors, mentre que, contràriament, temperatures de 80°C durant una hora augmenten els índexs de germinació. La calor pot no ser necessària per a la germinació, però l’estimula; el foc, com que destrueix les espècies llenyoses, elimina la font de la inhibició. Però, així que les espècies llenyoses s’instal·len novament, torna a aparèixer la inhibició de les llavors, les quals, tanmateix, es mantenen viables fins que es produeixi un altre incendi. Aquest fenomen d’al·lelopatia encara no ha estat constatat a la conca mediterrània, però hi ha una competició molt forta per la llum i l’espai, deguda a les plantes perennes, de manera que les plantes anuals o bianuals no poden persistir.
El cas extrem de la pirofília
El pirofitisme, és a dir, l’adaptació al foc que va més enllà de la simple capacitat de resistència, arriba a extrems molt acusats, ja que en el cas de determinats piròfits el foc ha esdevingut necessari per a la seva reproducció. Aquest és el cas de molts pins mediterranis i d’algunes bànksies (Banksia) australianes, els vegetals anomenats piròfils.
Els pins (Pinus) acumulen una gran quantitat de resines i essències volàtils, entre les quals hi ha la trementina, un producte fins fa molt poc obtingut exclusivament del resinat de determinades espècies de pins. Aquestes resines i essències es troben en totes les parts de la planta i fan que aquests arbres siguin terriblement inflamables, de manera que els incendis són molt més devastadors en les masses arbòries dominades per pins que en les dominades pels típics esclerofil·les mediterranis. Aquesta inflamabilitat, però, no va acompanyada de capacitat per a rebrotar; el foc provoca la mort d’aquests arbres i la regeneració queda limitada a la germinació. El problema associat a la freqüència massa alta d’incendis ha donat lloc a temps de maduració relativament baixos en els pins mediterranis, que presenten un període des de la germinació fins a la producció de llavors fèrtils més curt que el d’altres congèneres.
Certs pins posseeixen pinyes que només són dehiscents després del pas del foc o sota l’acció del foc. Són els que tenen pinyes seròtines, amb les bràctees unides per una resina o cera sensible a les altes temperatures, que es fon al pas de les flames i així permet que les pinyes s’obrin i deixin caure les llavors. Aquest fenomen ha estat molt estudiat a Amèrica del Nord, però també a la conca mediterrània, en alguns llocs, el pi blanc (Pinus halepensis) es podria comportar d’aquesta manera. Les pinyes, efectivament, actuen com a protectores de les llavors davant del foc. Després d’un incendi, les pinyes presenten un aspecte renegrit, però els pinyons que contenen no han patit l’efecte de les elevades temperatures. A més, l’escalfor contribueix a obrir les pinyes, tot alliberant una gran quantitat de llavors que colonitzaran el sòl nu amb relativa facilitat. Aquest comportament, que pot observar-se en espècies de la conca mediterrània com el pi blanc (Pinus halepensis), el pi de Calàbria (P. brutia), el pinastre (P. pinaster) i el pi pinyer (P. pinea), s’accentua en l’àrea californiana, on hi ha tota una colla d’espècies del mateix gènere (P. attenuata, P. muricata, P. radiata, P. remorata, P. torreyana, P. contorta) que només obren les pinyes per efecte de l’escalfor. En aquest cas, doncs, el foc esdevé necessari per a alliberar el potencial reproductiu emmagatzemat en les pinyes que es van acumulant any rere any en les capçades dels arbres. Els preciosos pinyons només cauran en terres convenientment netes de competidors.
Presenten un comportament semblant moltes espècies de proteàcies arbustives australianes, especialment les bànkies (Banksia). En aquest cas, les llavors queden tancades i emmagatzemades en fruits llenyosos i densos que recorden vagament les pinyes i que tenen exactament la mateixa funció (el cas més espectacular és el de la “bull banksia”, espècie que no solament es comporta de la manera descrita, sinó que, a més, és capaç de rebrotar. El foc és l’encarregat d’alliberar les llavors i, si no fa acte de presència, els fruits es van acumulant fins a constituir enormes reserves de material inflamable que acaben afavorint el foc. Les pinyes no s’obriran fins que no siguin sotmeses a l’escalfor del foc i les llavors no germinaran fins que el foc no n’hagi cremat parcialment la capa superficial protectora o estimulat canvis fisiològics interns. Moltes espècies d’aquests arbustos produeixen prodigioses quantitats de llavors: se n’han enregistrat mitjanes de deu milions per hectàrea i any, i no és estrany trobar més de mil llavors per metre quadrat a la capa superficial de sòl als boscos esclerofil·les humits de l’oest d’Austràlia. Després de l’incendi, les llavors alliberades germinen en el terreny buit i adobat per les cendres, mentre que els individus adults regeneren ràpidament la seva capçada tot rebrotant, no solament des de la soca, sinó també des de branques relativament primes que han quedat protegides per l’escorça.
En el cas dels eucaliptus (Eucalyptus), la inflamabilitat pot incrementar-se gràcies als olis volàtils de les fulles i a l’enorme producció de virosta eixarreïda. Efectivament, ultra grans quantitats de fulles, molts eucaliptus desprenen cada any plaques d’escorça seca, i d’altres tenen filagarses d’escorça seca que pengen de les branques, la qual cosa facilita que el foc es propagui del sotabosc a la capçada. Aquesta pirofília tan accentuada s’explica com un pas més del pirofitisme: la vegetació resistent al foc evoluciona fins a dependre’n per al seu manteniment i la seva regeneració.
A Victòria i a Tasmània, ja fora del bioma mediterrani, on els boscos esclerofil·les i hidrofítics conviuen l’un al costat de l’altre, les espècies dels boscos hidrofítics envairien de manera continuada els boscos d’eucaliptus en absència de foc, i finalment acabarien per desplaçar-los, si continués el procés durant el temps necessari (caldrien almenys 200 anys sense foc per a aconseguir-ho). En cas de produir-se un incendi, les espècies dels boscos hidrofítics serien eliminades i la successió hauria de tornar a començar. En els boscos esclerofil·les mediterranis es produirien fenòmens anàlegs, si mai no s’hi declarés cap foc. El foc, doncs, factor ambiental aparentment i realment tan devastador, ha estat internalitzat de tal manera en el cicle biològic d’alguns vegetals mediterranis, que ha esdevingut necessari per a la seva subsistència.
Les estratègies adaptatives
Les diferents respostes al foc de les espècies que es regeneren, sigui per via vegetativa sigui obligatòriament per via sexual, plantegen algunes preguntes. Sembla que els vegetals que tenen possibilitat de rebrotar de manera vigorosa presenten un avantatge considerable per a la recuperació de zones incendiades. Així que una espècie d’aquest tipus colonitza l’espai, l’ocupa durant molt de temps. Contràriament, el tipus de reproducció per via sexuada sembla clarament menys avantatjós perquè necessita instal·lar-se, distància de dispersió de les llavors i un índex de renovació de les plàntules.
Es fa difícil explicar la coexistència de dues vies tan diferents en els vegetals de les comunitats mediterrànies afectades pel foc, i hom es pregunta quins avantatges té per a una espècie utilitzar-ne una o altra. En particular, per a les que es reprodueixen obligatòriament per llavors, perquè, efectivament, pot produir-se un incendi que destrueixi tota la població abans que els individus hagin arribat a l’estadi adult i, per consegüent, siguin capaços de produir, novament, flors i llavors. L’interval de temps de descans entre dos incendis ha de ser ampli per a les espècies que només es reprodueixen per via sexuada, i pot ser més curt per a les dotades de regeneració vegetativa. Aparentment, un tipus (el vegetatiu) serveix per a la recuperació de l’espai prèviament ocupat, mentre que l’altre (el germinatiu) ofereix possibilitats d’envair les zones no ocupades prèviament. La coexistència de tots dos tipus deu provenir d’una alternança de pertorbacions i fases de repòs successives, que han condicionat la pressió de selecció que ha existit en el curs del temps.
Els efectes del foc sobre la vegetació i el sòl
La major o menor freqüència dels incendis afavoreix diferents estratègies i incideix, a més, en els processos de selecció i especiació. La superposició d’aquests processos dóna lloc a una més gran abundància d’espècies germinadores enfront de les rebrotadores, ja que el cicle vital més curt de les primeres permet un procés d’especiació més ràpid. Així, en dos gèneres típics del “chaparral”, Arctostaphylos i Ceanothus, hi ha respectivament 16 i 12 espècies que rebroten, enfront de 59 i 46 que germinen; i entre els típics del “fynbos” com ara Leucadendron i Leucospermum, les espècies rebrotadores són 7 i 4, enfront de 73 i 36 de germinadores. Aquesta relació d’abundància dins del tàxon és diferent quant a abundància i dominància en les comunitats, ja que la freqüència del foc és la que determina, en cada cas, quin tipus d’espècie i estratègia resulta més afavorida.
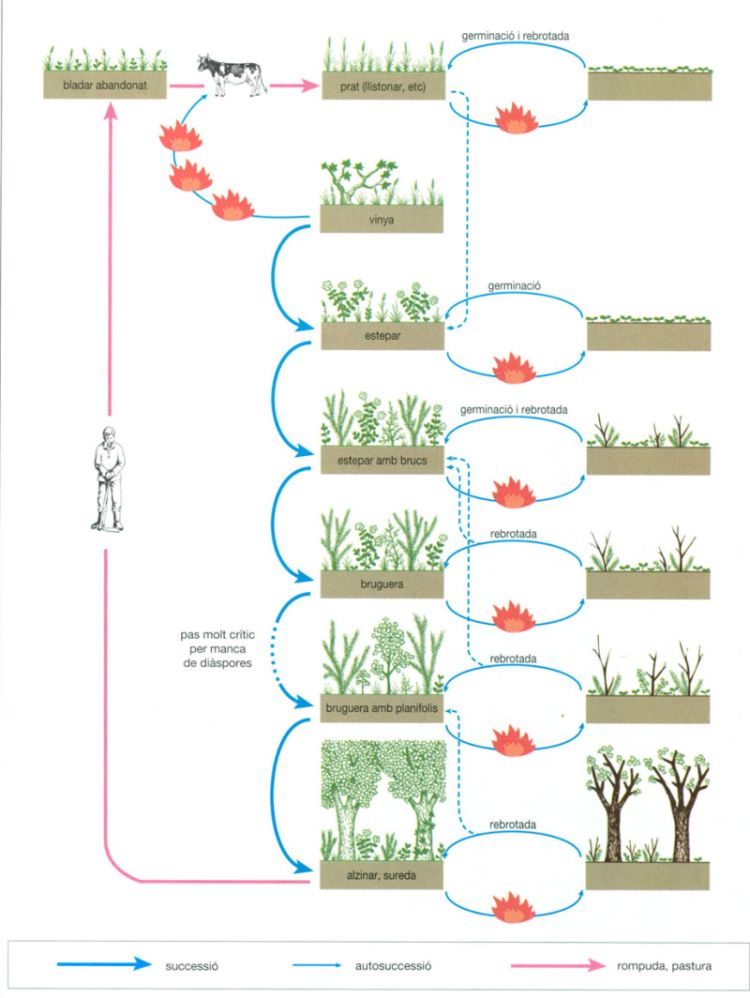
Models de successió de la vegetació després d’un incendi, al cap de Creus (NE de la Península Ibèrica). Si hom abandona una vinya —prèviament establerta, per exemple, en un espai forestal—, la successió restaura un bruguerar i fins i tot un bosc mediterrani, posat que hi hagi arbres llavorers a la rodalia; si bé l’escassesa o la manca de llavors pot fer crític el pas de recuperació de la vegetació autòctona. Després d’un foc, per contra, sol restaurar-se la mateixa comunitat que hi havia abans de la cremada, i el fenomen s’anomena d’autosuccessió.
Jordi Corbera
L’efecte conjunt de totes aquestes adaptacions dóna a la vegetació mediterrània una alta capacitat de regeneració després dels incendis: segons la climatologia, el creixement de la vegetació mediterrània després d’un incendi oscil·la entre 1 i 4 t/ha i any, una xifra gens menyspreable en aquest bioma i superior a la producció de formacions mediterrànies no afectades pel foc. El foc destrueix la vegetació i engega un procés anomenat d’autosuccessió, diferent al de la successió secundària. Aquest segon, que té lloc quan desapareix totalment la vegetació, incloent-hi les parts subterrànies i les llavors, és un llarg procés que porta de la terra nua fins a la vegetació climàcica del lloc mitjançant una successiva colonització i substitució d’espècies, des de les pioneres fins a les corresponents als estadis finals. Així, si un terreny ocupable per una màquia és roturat per al conreu i després s’abandona, s’engega un lent procés que conduirà a la regeneració de la màquia després de passar per diferents estadis. En canvi, si l’agent destructor és el foc, es regenera ràpidament el mateix tipus de vegetació a partir dels rebrots i les llavors de les mateixes espècies presents abans de l’incendi. La capacitat per a rebrotar fa que fins i tot els mateixos individus continuïn existint: en formacions arbustives baixes com el “chaparral” s’han trobat soques amb una edat estimada d’uns 250 anys, cosa que posa de manifest que un vegetal pluricentenari no ha de ser, necessàriament, un arbre esplendorós. La regeneració per autosuccessió en aquests boscos és rapidíssima, ja que són constituïts per arbres protegits per escorces gruixudes capaços d’emetre rebrots epicòrnics, per arbustos de talla mitjana que es comporten com a rebrotadors epicòrnics o de soca, i per arbustos de talla petita capaços de rebrotar de soca, en una combinació d’estratègies que es correspon amb la intensitat del foc en cadascun dels estrats del bosc.
En alguns casos, el foc sembla necessari per a mantenir la vegetació en un estat vigorós, ja que l’absència d’incendis dóna lloc a una decadència amb elevada acumulació de biomassa morta, cosa que afavoreix indirectament el foc; aquest és el cas dels paisatges californians dominats per “chaparral” i dels sud-africans i els australians dominats per proteàcies. En el cas dels boscos australians de “jarrah”, el foc sembla actuar com un element necessari per a mobilitzar periòdicament els nutrients immobilitzats en una virosta que es descompon molt lentament, tot accentuant la pobresa en nutrients del sòl. Les condicions climàtiques imperants a Austràlia fan que aquests detrits es descomponguin molt lentament i continuïn acumulant-se durant 30-40 anys abans que s’arribi a un equilibri entre acreció i descomposició. Aquesta virosta s’acumula al sòl del bosc a un ritme que se situa entre 1 i 3,5 tones per hectàrea i any en el bosc esclerofil·le sec, i entre 2,5 i 11 tones en l’esclerofil·le humit.
Però l’elevada capacitat de regeneració després del foc que té la vegetació esclerofil·la mediterrània no comporta pas una immunitat absoluta per al bosc, ja que els incendis perjudiquen la fauna i, singularment, el sòl. En efecte, el factor negatiu més important associat als incendis és l’erosió, molt acusada quan les pluges torrencials típiques del clima mediterrani incideixen sobre sòls desprotegits de la coberta vegetal. Cal remarcar que la regeneració posterior a un incendi pot ser molt lenta durant el període eixut estival, cosa que fa que les primeres i violentes pluges tardorals actuïn sobre sòls nus. L’erosió representa una pèrdua gairebé irreversible sota unes condicions ambientals que donen lloc a una edafogènesi lentíssima o nul·la, i compromet la futura regeneració de la vegetació. En zones de “chaparral” s’ha comprovat que l’erosió, que provoca la pèrdua d’un gruix de sòl de fins a 7 cm, es veu incrementada fins a vuit anys després de l’incendi. Aquest increment de l’erosió es deriva de la pèrdua de coberta vegetal i de la destrucció de l’estructura edàfica a causa de les altes temperatures, ja que en un incendi que assoleix en superfície els 1 100°C, els primers centímetres del sòl poden arribar a temperatures de 250°C, cosa que provoca una destrucció de tota la matèria orgànica d’aquestes capes superficials, amb la consegüent pèrdua de coherència.
És interessant fer notar que el problema de la destrucció del sòl i de la pèrdua de nutrients associat als incendis es fa especialment greu quan la comunitat cremada és arbustiva, tal com succeeix molt sovint en el bioma mediterrani, ja que l’acumulació de biomassa prop del sòl fa que pugi molt la temperatura en els seus primers nivells. En un incendi de “chaparral” en què s’arriba a temperatures de prop de 1 000°C en les capçades, la superfície del sòl pateix temperatures de 700°C, mentre que en un incendi d’un bosc de coníferes, la mateixa temperatura en capçades més altes provoca un escalfament del sòl fins a 300°C, i en el cas d’un prat, amb menys massa de combustible, el sòl no s’escalfa més de 250°C. Aquestes elevades temperatures provoquen, a més, una volatilització de nutrients essencials com el nitrogen, tant de les primeres capes del sòl com de la biomassa, i donen lloc a un empobriment progressiu molt greu, ja que sovint es parteix de sòls marcadament pobres.
Evidentment, l’elevada freqüència d’incendis derivada de l’activitat humana accentua el problema de l’erosió i de l’empobriment edàfic, i treu efectivitat a algunes de les adaptacions abans esmentades. Les espècies amb capacitat per a rebrotar també pateixen els problemes derivats dels incendis recurrents, ja que cada soca pot rebrotar un nombre limitat de vegades. A més, sobre sòls magres i en condicions de secada forta, un percentatge molt elevat (fins al 50%) dels individus amb capacitat per a rebrotar moren després de l’incendi. El foc, un vell conegut del bioma mediterrani, ha esdevingut, de la mà dels humans, un important factor de degradació paisatgística, tal com serà analitzat més endavant.
El foc i el cicle dels nutrients
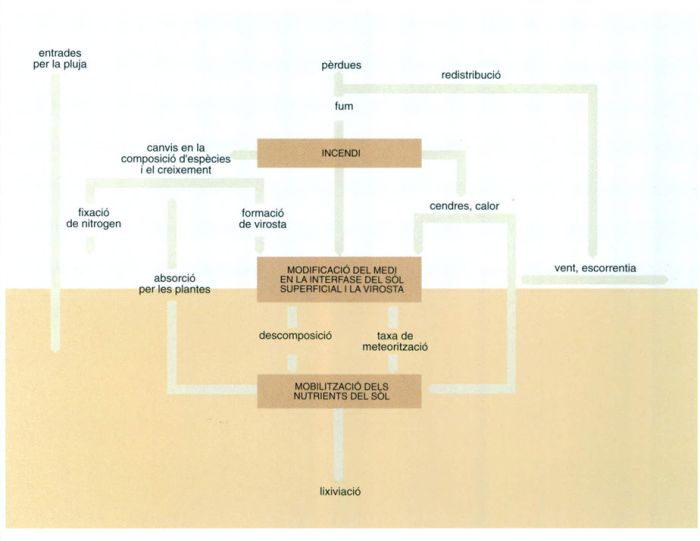
El foc té efectes sobre el cicle dels nutrients, ja que mobilitza els que hi ha continguts en els vegetals vius i els diposita en forma de cendres en la mateixa contrada. Alguns d’aquests nutrients, però, es perden amb el fum del foc, i el fet que les cendres es dipositin sobre un sòl despullat de vegetació afavoreix que la pluja o el vent posteriors a un incendi agreugin encara més la pèrdua de nutrients en ésser traslladats a altres ecosistemes, veïns o llunyans.
Editrònica
Els ecosistemes de les regions de clima mediterrani són considerats, generalment, ecosistemes amb pocs nutrients, si bé hi ha diferències en la quantitat de reserves del sòl entre les cinc àrees amb un clima de tipus mediterrani. Durant un incendi i després, el subsistema constituït per l’humus del sòl és directament i indirectament afectat per l’acció del foc. Com que la fertilitat del sòl és el factor crític, que controla l’estructura i la funció de les comunitats vegetals i dels ecosistemes en conjunt, és important comprendre clarament els efectes del foc en les reserves de nutrients.
Els nutrients són emmagatzemats en quantitats variables a la vegetació, a la capa d’humus i al sòl. Les reserves de nutrients de la vegetació depenen bàsicament de l’edat del bosc, la biomassa aèria i la concentració de nutrients; també són influïdes per la distribució relativa del fullatge en la biomassa, perquè les fulles generalment contenen més nutrients que els teixits llenyosos. Com més antic és un bosc, més abundant és el compartiment llenyós i més gran el risc d’incendi. Les reserves de nutrients de l’humus es troben a la capa on s’acumula el material orgànic mort, parcialment o totalment descompost. La composició elemental d’aquest material depèn de la forma de deposició de l’humus quan cau al sòl, de la velocitat d’acumulació i descomposició —fortament influïda per la composició química—, del volum del compartiment foliar comparat amb el llenyós i de les possibles pèrdues a causa de l’erosió i de la lixiviació. Les reserves de nutrients solubles en aigua (potassi, calci i magnesi) estan fortament influïdes pel cicle hidrològic. Finalment, les reserves de nutrients del sòl depenen del tipus de roca mare local, del patró climatològic, de l’edat del sòl i de la vegetació i, naturalment, dels factors antropogènics que actuen sobre el sòl. En general, els sòls dels ecosistemes mediterranis són deficients en nutrients, especialment en nitrogen i fòsfor.
Els mecanismes pels quals el foc afecta les reserves i el cicle dels nutrients són bastant complicats. El foc mobilitza els nutrients incinerant la biomassa del bosc i la matèria orgànica de l’humus i del sòl, de manera que el resultat és la deposició de cendres. Hi ha alguns nutrients que es perden amb el fum del foc, i n’augmenta la pèrdua potencial per la pluja i/o l’acció del vent (lixiviació, escorrentia i acció eòlica). Alguns nutrients transferits a l’atmosfera es dipositen al mateix ecosistema, un cop el foc és extingit, mentre que els altres ho poden fer prop del lloc cremat o bé molt lluny.
A aquestes respostes abiòtiques caldria afegir els efectes produïts per processos biològics induïts o augmentats pel foc. En altres paraules, els efectes del foc en el cicle i en les reserves de nutrients depenen en gran manera dels processos específics de cada ecosistema, que impliquen interaccions entre les condicions climàtiques prevalents, la condició del sòl i la vegetació i les interferències humanes.
Els incendis de la vegetació tenen múltiples efectes en les reserves de nutrients del sistema sòl-plantes i, en conseqüència, en els cicles de nutrients dins de l’ecosistema. En general, el foc és considerat un ràpid descomponedor, perquè determina la majoria dels límits de nutrients de la biomassa del bosc o de l’humus disponibles per a les plantes. De tota manera, aquesta conseqüència ecològica actua a favor de l’ecosistema només sota certes condicions relativament ben definides d’intervals normals entre incendis.
Sovint els incendis donen com a resultat un esgotament dels nutrients i una desertització de l’àrea cremada, pels efectes acumulatius sobre les reserves de nutrients. Però si l’interval natural entre incendis (considerat entre 25 i 30 anys per la majoria de científics) es manté, aquest risc ambiental pot ser considerat perfectament un agent que promou el rejoveniment i l’estabilització dels ecosistemes.
Els efectes sobre el nitrogen
Entre els nutrients més importants, el nitrogen mereix un tractament especial per les particulars propietats químiques i l’escassetat al sòl dels ecosistemes mediterranis. Per l’acció directa del foc, el nitrogen és en gran part volatilitzat per la potència calorífica durant un incendi. Es considera que de l’energia alliberada en forma de calor durant un incendi, només el 8% és absorbida i transferida a les capes del sòl. Les pèrdues potencials de nitrogen estan fortament relacionades amb les temperatures assolides durant un incendi. Quasi no hi ha pèrdues detectables a temperatures per sota dels 200°C, mentre que les pèrdues més grans observades es donen als 300-400°C (75-100%); per sobre dels 500°C es podrien perdre pràcticament totes les reserves de nitrogen de les plantes i l’humus. Les pèrdues de nitrogen del sòl depenen principalment del contingut d’humitat. L’aigua té una elevada capacitat calorífica i una elevada calor de vaporització, que impedeixen que el sòl s’escalfi a temperatures per sobre dels 100°C mentre l’aigua no està totalment evaporada o és transferida a capes més profundes. La textura del sòl també té un paper important en la transferència de calor durant un incendi.
Tot i que la major part del nitrogen es volatilitza durant un incendi, les formes disponibles d’aquest element es troben en més quantitat en els llocs cremats que en els que no ho estan, a causa de la ràpida descomposició i mineralització de l’humus i del subsegüent enriquiment del sòl després d’un incendi. En general, després d’un incendi es troben concentracions més elevades d’amoni-nitrogen (NH+4-N), mentre que la velocitat de nitrificació augmenta remarcablement. Fins i tot si s’accepta que immediatament després d’un incendi la quantitat total de nitrogen es redueix, les condicions edàfiques després del foc afavoreixen i augmenten enormement l’activitat microbiana del sòl. La fixació de nitrogen per microorganismes de vida lliure és promoguda pel foc i es considera induïda pel canvi de pH del sòl cap a valors més neutres.
A més de la fixació de nitrogen per part dels microorganismes de vida lliure, hi ha proves que també es dóna una fixació per part d’organismes simbionts. Això s’atribueix a la proliferació de plantes lleguminoses durant les etapes immediatament posteriors a l’incendi. La família de les lleguminoses —les fanerògames ecològicament més reeixides del món— és la primera en els matollars mediterranis pel que fa al nombre de gèneres (15 de cada 76, o el 19,7%), i constitueix aproximadament el 8% de la flora dels països de la conca mediterrània. De tota manera, aquest nombre pot ser subestimat, perquè s’hi inclouen alguns països que al costat del bioclima mediterrani en tenen d’altres, com per exemple Algèria, Síria, Egipte, o el nord de França. Els nombres relatius per a altres regions del món amb clima de tipus mediterrani varien entre el 3,8% i el 13,3%, per a Xile i el sud-oest d’Austràlia respectivament. Les lleguminoses han estat considerades un component important de les comunitats vegetals que prosperen després d’un incendi als ecosistemes mediterranis.
Segons la informació de què disposem fins ara, és evident que hi ha un enriquiment de les flores locals en lleguminoses. Aquest enriquiment pot ser lleuger, com és el cas del “chaparral” de Califòrnia, on la flora autòctona de lleguminoses augmenta només el 7,3%, o realment impressionant, com en llocs cremats de Grècia, on s’ha observat un augment superior al 50%, amb una gamma intermèdia de respostes. La proliferació de plantes lleguminoses a les àrees cremades només es dóna durant les etapes immediatament posteriors a l’incendi. La majoria d’aquestes espècies són plantes anuals o, com a molt, biennals, que pertanyen als gèneres Vicia, Lathyrus, Lotus, Medicago i Trifolium, i que, gradualment, es van restringint en nombre d’espècies i en cobertura, formant un banc de llavors que espera el pròxim incendi. La fixació de nitrogen per les lleguminoses herbàcies de vida curta encara s’està investigant. De tota manera, la quantitat de nitrogen que retorna al subsistema del sòl en forma d’humus a punt de descompondre’s hauria de ser considerada una font addicional important d’aquest element.
Els efectes sobre el fòsfor
Sembla que el fòsfor no resulta tan afectat pel foc; com a mínim tots els autors hi estan d’acord. Al contrari, les formes disponibles d’aquest element augmenten després d’un incendi, fet d’extremada importància si es té en compte que la vida del fòsfor al sòl dels ecosistemes mediterranis és molt curta.
Els efectes sobre els cations
Bé que s’han considerat algunes pèrdues de cations, la disponibilitat de la majoria s’incrementa immediatament després d’un incendi. En les cendres dipositades sobre el sòl cremat s’han mesurat quantitats relativament altes de potassi, calci i magnesi. Això no obstant, l’increment de la disponibilitat d’aquests cations, com el potassi, que són relativament més mòbils, es redueix en l’etapa de pre-foc de les àrees no cremades, possiblement a causa de la lixiviació. Molt sovint s’observa un canvi en el pH del sòl com a resultat dels cations alliberats a les cendres per la combustió de les plantes i de la matèria orgànica del sòl. L’ordre d’aquest canvi i el temps que dura són funció de diferents paràmetres, entre els quals tenen una importància cabdal el pH original, la quantitat de cendres produïdes, la composició química i el règim de precipitacions.
Els efectes sobre l’activitat microbiana del sòl
El canvi de pH del sòl després d’un incendi afecta directament o indirectament l’activitat microbiana. En les poblacions bacterianes, remarcablement activades, s’observa una resposta immediata. A molts llocs de la regió mediterrània s’ha descrit un augment de la velocitat d’amonificació i de nitrificació després d’un incendi. Els fongs sembla que inicialment queden afectats desfavorablement, perquè en general no es desenvolupen bé en pH àcids. Els processos de fixació de nitrogen per microorganismes de vida lliure i simbionts ja han estat descrits més amunt. Se sap molt poc sobre l’efecte del foc en els propàguls de les micorizes en les condicions edàfiques posteriors a un incendi. Se suposa que, com passa amb altres grups de fongs, els que formen associacions de micorizes resulten afectats negativament pel foc, sobretot a causa del canvi de pH cap a valors més àcids, però també a causa de l’increment de temperatura del sòl, que, de tota manera, rarament arriba a valors letals. Els mitjans emprats, la velocitat i el grau de recobriment de les associacions de micorizes de les arrels de la majoria de plantes de les regions mediterrànies són molt importants, perquè acompleixen un paper bàsic en l’establiment i la supervivència de les plantes sota condicions edàfiques adverses, ja que actuen com a arrels especialitzades que es ramifiquen a través de grans volums de sòl i traspassen els nutrients a les plantes a una velocitat molt més gran que les arrels no micorizades.
El cicle anual
L’activitat de la vegetació resulta condicionada per la variació que presenten al llarg de l’any els recursos que necessita, és a dir, la llum, l’aigua i els nutrients. Aquestes variacions estacionals donen lloc a una adaptació de l’activitat dels organismes, que poden utilitzar com a indicador directe el nivell d’un determinat recurs (per exemple, la humitat del sòl) o bé fer-ne servir un indicador indirecte (per exemple, la variació de durada del dia com a indicador de l’arribada del període fred o càlid). De tota manera, els organismes procuren desencadenar cada activitat en el moment en què poden disposar dels recursos necessaris. En el cas del clima mediterrani, hi ha una clara fluctuació estacional de la temperatura, de la humitat i de la disponibilitat de nutrients, que depèn directament de la disponibilitat d’aigua.
La temperatura presenta una fluctuació molt predicible i regular, i actua com a limitador per al creixement durant una part del període fred. La precipitació, en canvi, és poc regular i predicible, tot i que té tendència a acumular-se fora de l’estació càlida. La vegetació mediterrània, doncs, veu limitada la seva activitat per les relativament baixes temperatures hivernals i per la manca d’aigua i nutrients durant el període eixut, i concentra la seva activitat al llarg de la tardor i la primavera; els factors desencadenants són la disponibilitat d’aigua i la temperatura. Aquest esquema general admet, però, moltes matisacions, ja que les espècies han desenvolupat diferents mecanismes per fer front als mateixos problemes, cosa que permet l’activitat d’algunes quan d’altres romanen inactives.
Cal esmentar una clara diferència entre els comportaments generals de la vegetació en les zones de la conca mediterrània, Califòrnia i Xile, per una banda, i les d’Austràlia i Sud-àfrica per l’altra. Les primeres responen a l’esquema esmentat, i en les segones trobem un seguit d’adaptacions derivades de la marcada pobresa en nutrients de la majoria dels seus sòls. Això dóna lloc a una absorció i una acumulació de nutrients durant l’època més humida que són emprats per al creixement dels nous brots durant l’època càlida, o sigui, a ple estiu. Aquesta estratègia respon a la necessitat d’evitar la pèrdua de nutrients que són alliberats per descomposició precisament quan la humitat edàfica és més elevada, però les temperatures no són les òptimes per al creixement. De tota manera, la presència de plantes amb un ritme de creixement estival es relaciona, també, amb les influències tropicals en la gènesi de les flores mediterrànies de les regions australiana i capenca.
La brotada i el creixement vegetatiu
En termes generals podem dir que la brotada es concentra al llarg de la primavera, moment en què les temperatures elevades i la bona disponibilitat d’aigua i nutrients permeten un creixement actiu. De tota manera, hi ha dues estratègies clarament diferenciades: la dels esclerofil·les perennifolis i la dels malacofil·les caducifolis estivals, abundants aquests últims en les zones més càlides i seques. Els perennifolis, normalment d’arrelada profunda, tenen aigua a la seva disposició a partir del final de la tardor, quan les primeres pluges s’han infiltrat fins a capes profundes, però difereixen la brotada fins a la primavera per evitar els possibles danys produïts en els nous brots per les glaçades hivernals. Els caducifolis estivals, en canvi, d’arrelada més superficial i situats en els llocs més càlids, comencen la brotada al final de la tardor i mantenen el seu creixement fins a la meitat de la primavera, moment en què l’aigua comença a ser escassa. La plasticitat d’una vegetació adaptada a viure en un clima poc regular és, però, considerable, i en anys d’estius anormalment plujosos o d’hiverns anormalment suaus poden donar-se brotades addicionals o fora d’època per aprofitar les condicions favorables. Les plantes herbàcies i anuals poden presentar dues èpoques de creixement actiu, durant la tardor i la primavera, tot aprofitant els moments en què la combinació de temperatura i humitat és òptima.
El creixement en gruix dels troncs i de les arrels, com també l’allargament d’aquestes últimes, pot tenir lloc en altres moments de l’any a causa de la menor sensibilitat dels nous teixits formats a les baixes temperatures. Cal tenir present que l’activitat fotosintetitzadora es manté durant una bona part de l’any, subministrant recursos necessaris per a dur a terme aquestes activitats. Així, en termes generals, el creixement de les arrels s’activa durant la tardor mentre que l’engruiximent dels troncs es fa màxim en ple hivern. Aquí també apareixen totes les matisacions i les irregularitats possibles, de manera que en els vegetals llenyosos mediterranis es fa molt difícil el reconeixement d’anells de creixement i, en el cas que es reconeguin, és molt difícil saber si un anell correspon a un creixement anual o només a una part, a causa de la plasticitat de resposta. La informació climàtica acumulada en les seqüències de creixement dels vegetals llenyosos, tan útil i clara sota climes d’estacionalitat marcada, esdevé difícilment interpretable en el bioma mediterrani, en el qual arbres i arbustos escriuen la seva història amb una cal·ligrafia confusa.
La floració
Variació del percentatge de floració mensual de 18 espècies de plantes llenyoses a Echo Valley (San Diego, Califòrnia) i de 20 espècies de Fundo Santa Laura (Santiago, Xile) al llarg d’uns quatre anys. Tot i que les condicions climàtiques i de nutrients del sòl són similars a ambdues regions mediterrànies, hi ha diferència en la fenologia de les plantes: la distribució més uniforme de la floració al llarg de tot l’any en el cas de la regió xilena s’atribueix a la major diversitat de formes florístiques i al clima més moderat.
Editrònica
Les pautes de floració presenten una irregularitat encara més marcada, ja que la floració no és un procés que necessiti quantitats importants de recursos hídrics. Algunes espècies floreixen utilitzant recursos fabricats durant la mateixa estació de creixement, i d’altres fan servir reserves acumulades durant l’estació anterior, separant així la floració del període favorable per al creixement. Sembla que la floració està més relacionada amb la presència i la disponibilitat de pol·linitzadors que amb la d’aigua i nutrients. Així, la intensitat de floració d’algunes espècies arbustives, com per exemple el “chamizo” de Califòrnia (Adenostoma fasciculatum), està molt relacionada amb la disponibilitat d’aigua just abans de la floració, ja que fan servir recursos sintetitzats en aquell període. Altres de molt emparentades, com per exemple A. glauca, presenten una alta correlació entre intensitat de floració i disponibilitat d’aigua durant l’estació de creixement de l’any anterior, ja que desenvolupen els primordis florals en acabar el creixement i floreixen mesos després mobilitzant recursos de l’any anterior. Això dóna lloc a floracions esgraonades al llarg de pràcticament tot l’any, amb mínims estivals i hivernals.
A la part occidental de la conca mediterrània, per exemple, hi ha espècies que inicien la floració quan el dia s’escurça, és a dir, durant la tardor. Aquest és el cas de l’olivarda (Inula viscosa), un arbust caducifoli hivernal típic de les rambles i les rieres, que ha colonitzat molts conreus abandonats, coberts de flors grogues quan arriben les primeres pluges després de l’estiu. També floreix a la tardor un arbust perennifoli típic d’alzinars i màquies com l’arboç (Arbutus unedo), que queda cobert dels raïms de flors blanques al mateix temps que acaben de madurar els fruits produïts en la floració de l’any anterior.
En ple hivern, assistim a la florida de moltes plantes herbàcies arvenses que abunden en els conreus arboris de secà, com els oliverars, els garroferars i els tarongerars. Les més abundants són les ravenisses (Diplotaxis erucoides, Erucastrum nasturtiifolium) i el pa de cucut (Oxalis pes-caprae), que cobreixen de blanc i groc els espais entre els arbres.
Altres espècies herbàcies no arvenses, com els caps-blancs (Alyssum maritimum) floreixen també a l’hivern, fent companyia a alguns arbustos com la gatosa (Ulex parviflorus), el marfull (Viburnum tinus), el romaní (Rosmarinus officinalis) i les farigoles (Thymus). Algunes espècies introduïdes, com ara l’ametller (Prunus dulcis) i la mimosa (Acacia dealbata) acaben de donar la nota de color al paisatge durant l’època freda.
Quan arriba la primavera es produeix l’esclat més gran de floració, ja que entre el març i el juny floreixen la gran majoria de les espècies mediterrànies. Algunes comunitats, com les brolles d’estepes (Cistus), canvien radicalment d’aspecte i color a causa de l’aparició massiva de flors de mida considerable, mentre que altres, com els alzinars, només adquireixen una tonalitat lleugerament diferent, amb les capçades cobertes de flors d’un color groc verdós discret.
L’estiu torna a ser una època de floracions més aviat escasses però no pas absents. És el moment de plantes crasses, algunes introduïdes, com l’atzavara (Agave americana) i la figuera de moro (Opuntia ficus-indica). La floració estival és, però, més discreta que no pas l’hivernal, afectada per la manca d’aigua.
En els sistemes mediterranis, en definitiva, l’encavalcament de l’activitat de diferents espècies dóna lloc a una activitat general gairebé continuada, que no s’atura ni tan sols durant l’època eixuta en els llocs més àrids, ja que s’hi mantenen en activitat les plantes suculentes. La coexistència de diferents estratègies biològiques per a fer front a problemes semblants és la que explica aquesta situació.