Els orígens de la fauna mediterrània
Atès que l’origen del clima mediterrani és recent, les relacions de parentiu entre la fauna que ocupa actualment les cinc regions amb aquest tipus de clima s’han de cercar en esdeveniments anteriors, i especialment en la situació de les terres sorgides en el passat i en la major o menor proximitat d’aquestes regions en altres èpoques geològiques. La història d’aquestes relacions pot començar en el període juràssic de l’era mesozoica, amb la fragmentació del supercontinent que abans reunia gairebé totes les terres.
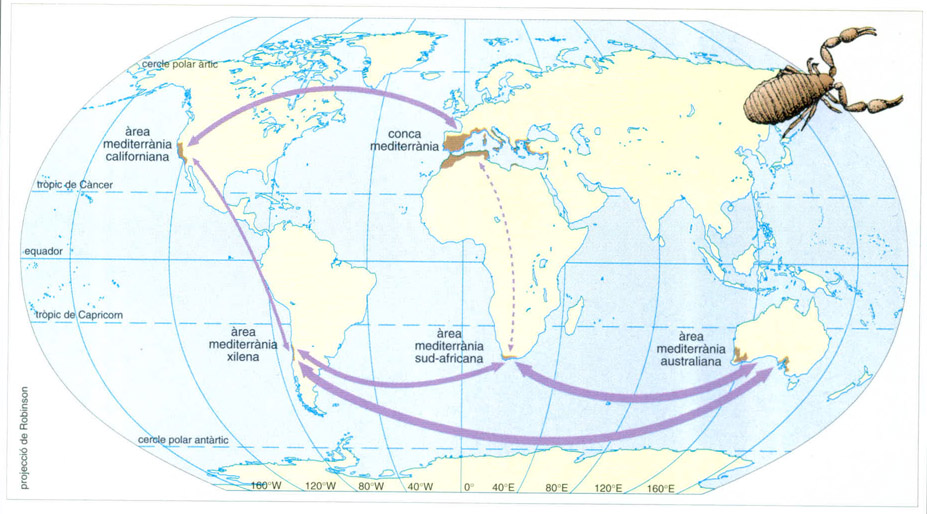
L’intercanvi de grups comuns fou possible entre Austràlia i Sud-amèrica fins a la darreria del Cretaci, a través de l’Antàrtida, que llavors es trobava unida a aquests continents, i entre Àfrica i Sud-amèrica fins al començament del mateix període; a l’hemisferi nord, l’intercanvi fou possible fins ben entrada l’era terciària, ja que la separació entre Amèrica del Nord i Euràsia fou tardana. Com que el clima mediterrani no aparegué fins a la darreria de l’era terciària, en què els continents i les àrees que serien mediterrànies ocupaven llurs posicions actuals, aquelles característiques comunes de la fauna que poden interpretar-se com a adaptacions no s’expliquen per raons biogeogràfiques, sinó més aviat per les restriccions imposades recentment per aquest tipus de clima. Tanmateix, en un cas, entre Califòrnia i Xile, hi hagué, a partir del Pliocè final, una connexió que permeté la migració de certs animals.
Les primeres manifestacions clares del règim climàtic mediterrani es troben al Plistocè, després de la primera glaciació. Davant l’aparició de la sequera estiuenca, la fauna hi respongué de diferents maneres, que configuren les estratègies de supervivència. Alguns grups foren simplement eliminats i només romanen en alguns enclavaments reduïts, on es conserven fins a cert punt les condicions del Cenozoic. Altres grups persistiren sense problemes, ja que es trobaven pre-adaptats a la sequera estacional, encara que llavors aquesta no tingués causes climàtiques. Aquests organismes serien fàcilment seleccionats en un ambient que limitaria la capacitat de competència de molts d’altres. Un tercer grup d’organismes es refugià en indrets diferents d’aquells on habitaven, a la recerca d’unes condicions estables que ja no podien trobar en llur medi habitual; així, molts elements de la fauna del sòl es traslladaren simplement cap a capes més pregones o esdevingueren habitants de coves. D’altres cercaren refugi en indrets amb oscil·lacions climàtiques mitigades, com ara pendents arrecerats, congostos o brolladors freàtics. Alguns endemismes mediterranis, com ara el ferreret (Alytes muletensis), han sobreviscut probablement gràcies a aquest recurs. Per últim, alguns grups arribaren a evolucionar veritablement en el breu espai de temps disponible i a adaptar-se a les noves condicions.
L’efecte de les glaciacions
Fa uns tres milions d’anys s’inicià el règim tèrmic oscil·lant que ha caracteritzat el final del Pliocè i tot el Plistocè. Malgrat que hom coneix pocs detalls de les seves manifestacions més antigues, hom suposa que el nombre total d’oscil·lacions, amb el consegüent avenç i retrocés dels glaços polars, és de 16 a 25. Les glaciacions finals, més ben conegudes, són les que tingueren lloc al Plistocè, durant el darrer milió d’anys. Per a la fauna mediterrània, aquestes glaciacions comportaren canvis d’importància fonamental, segons la regió considerada. A Austràlia i a Sud-àfrica tingueren poca transcendència, ja que els animals disposaven d’un ampli espai per a arrecerar-se del fred, i no hi ha muntanyes prou elevades perquè s’hi pogués acumular glaç.
El cas de Califòrnia, i sobretot de Xile, és diferent: allí sí que hi ha muntanyes altes, on s’acumularen masses de glaç més o menys extenses, que es retiraven durant els períodes interglacials. La fauna es veia obligada a allunyar-se de les muntanyes i a anar cap a la costa o cap a l’interior, i a ocupar els fons de les valls arrecerades i els refugis costaners. Algunes d’aquestes valls i aquests refugis quedaven envoltats de zones més fredes i, aïllats d’aquesta manera, els animals termòfils del bioma mediterrani podien evolucionar d’una manera independent durant unes quantes desenes de milers d’anys. La situació s’invertia en els períodes interglacials: les espècies termòfiles avançaven seguint els glaços en retirada, i les espècies estenotermes romanien aïllades en nuclis elevats, propers als cims. Aleshores, els processos de diferenciació i especialització tenien lloc per a aquestes últimes. Per tant, hom ha pogut parlar de dos tipus d’especiació a la regió central de Xile, l’especiació de vall i la de cim.
A Califòrnia el patró era similar: també hi havia els refugis costaners i de vall durant els períodes glacials, però com que les muntanyes de Sierra Nevada són més baixes que els Andes, les masses de glaç que s’hi acumulaven eren menors i el grau d’aïllament i d’especialització, també menor. D’altra banda, el casquet glacial d’Amèrica del Nord era molt gran, i en el seu avenç imposava una direcció general de retirada de les espècies cap al sud, que predominava sobre l’efecte de les muntanyes. En els períodes interglacials s’invertia el sentit d’avenç, si bé la durada d’aquests períodes ha estat molt més breu que la dels períodes freds; però en tot el procés, les espècies es podien retirar amb facilitat a zones més calentes (o més fredes), cosa que permetia d’evitar-ne l’extinció. Tot això garantí la permanència d’un gran nombre d’espècies a Amèrica del Nord, on la diversitat d’arbres i de mamífers és molt més alta que en regions comparables d’Europa.
El cas de les àrees que circueixen la mar Mediterrània és molt especial. A Europa hi hagué, diverses vegades, un extens casquet de glaç que empenyia, en els seus avenços, la fauna i la flora termòfiles cap al sud, i també hi hagué casquets glacials secundaris als Alps i en altres serralades muntanyoses. Ara bé, aquestes cadenes tenien preferentment una direcció E-W i podien constituir veritables barreres que impedien el trasllat lent cap al sud. Per això, potser, moltes espècies desaparegueren en les diferents glaciacions, i encara avui la riquesa específica a Europa és baixa per a diversos grups de plantes i animals.
Però això no passà en altres grups; als extrems orientals i occidentals dels Pirineus, els Alps i els Càrpats hi continuava havent corredors, on les condicions climàtiques eren més suaus, i pels quals corria el flux i el reflux de les espècies de l’una i l’altra banda. Les espècies termòfiles assolien, d’aquesta manera, les penínsules mediterrànies, on podien prosperar a redós d’un clima menys dur. Així, parcialment aïllades de llurs congèneres continentals —fins i tot durant els breus interglacials—, podien diferenciar-se i desenvolupar característiques peculiars.
El paper dels refugis càlids perimediterranis en l’evolució de les faunes locals ha estat molt estudiat. Aquests refugis actuaren com a centres de dispersió, des dels quals s’estengueren les espècies i les formes que hi aparegueren, tal com ho feren en altres centres de dispersió de l’hemisferi nord. Precisament el sud de Califòrnia, una altra zona de clima mediterrani, també és un d’ells; però la conca mediterrània és el més gran i el més conegut. A llur torn, es dividien en refugis menors, on es concentraven poblacions o formes diferents. Per exemple, al centre de dispersió mediterrani hom pot distingir nou unitats que funcionaren com a refugis independents; la més gran d’aquestes unitats, anomenada refugi atlanto-mediterrani, era una zona que abraçava la Península Ibèrica i el nord del Marroc i d’Algèria. Altres subrefugis importants foren el balcànico-anatòlic, l’itàlic i el mauritànic, aquest darrer entre el Sàhara i l’Atles. Com que les penínsules mediterrànies tenen un interior muntanyós, probablement els animals més termòfils s’acumularen en àrees reduïdes a la vora de la costa.
L’especiació insular
Les glaciacions foren acompanyades d’un moviment simultani de retirada i avenç del mar. La fosa dels glaços septentrionals augmentava el nivell del mar, amb la qual cosa els punts elevats i els turons pròxims a la costa podien quedar aïllats i formar illes durant alguns milers d’anys. Ben al contrari, amb l’avenç de la glaciació, el mar es retirava i les illes pròximes a la costa hi quedaven unides tot formant tomboli i penínsules. Com que les espècies termòfiles llavors eren acantonades en refugis costaners, era fàcil que ocupessin aquestes penínsules i que, en retirar-se els glaços, es formessin poblacions separades de la resta de l’espècie a les illes resultants. Això donà lloc a un procés repetit de diferenciació i formació de cercles de races, que es mostra especialment palès a les illes de la Mediterrània. Com calia esperar, atès l’origen d’aquestes poblacions diferenciades, sovintegen més en espècies terrestres i termòfiles, que depenguin més de les condicions climàtiques i que no tinguin capacitat de desplaçar-se ràpidament a grans distàncies fugint de circumstàncies adverses. En resum, el procés de microevolució insular és característic dels petits rèptils mediterranis i d’altres grups com els coleòpters o mol·luscs terrestres, que presenten també les condicions esmentades.
La majoria de les espècies de rèptils mediterranis de mida petita queden incloses en la família dels lacèrtids. Un gènere de sargantanes, Podarcis, ha estat, potser, el protagonista més destacat dels esmentats processos de formació de races i subspècies insulars.
A l’Europa central i occidental hi ha una espècie d’aquest gènere, Podarcis muralis, que es pot considerar pròxima a l’estoc que hi havia al continent abans del Plistocè. Les glaciacions successives obligaren aquest estoc a acantonar-se a les diferents penínsules mediterrànies, on evolucionà independentment i donà lloc a diferents espècies: Podarcis hispanica, per a la Península Ibèrica, P. sicula, per a la Itàlica, i P. erhardii, per a la Balcànica. La diferenciació d’aquestes espècies s’inicià ja al Pliocè, ha prosseguit fins avui i ha donat lloc a altres espècies pròximes, tant a les penínsules esmentades (per exemple, P. bocagei) com a les illes mediterrànies (per exemple, P. lilfordi, P. tiliguerta o P. filfolensis).
Moltes d’aquestes espècies ocupen una àrea continental o insular voltada d’illots menors pròxims. El fenomen d’aïllament i comunicació alternants s’ha repetit per a aquests illots amb més freqüència que no pas per a les illes allunyades del continent, atès que la profunditat dels braços de mar que les separen de la terra és baixa i, per tant, n’hi ha prou amb oscil·lacions climàtiques menors per a alterar-ne el grau de connexió amb aquesta. D’aquí que els arxipèlags mediterranis que posseeixen molts illots tinguin, també, una gran varietat de formes locals de sargantanes, generalment amb trets morfològics propis i que s’adscriuen a un o a uns quants d’aquests illots.
El cas més notable és el de la sargantana pitiúsica (Podarcis pityusensis), que viu a les illes Balears, a Eivissa, Formentera i els illots pròxims. Hom ha descrit 42 subspècies d’aquesta espècie, en general corresponents a les poblacions de cada illot. Actualment, hom tendeix a reduir-ne el nombre, però en tot cas és evident que hi ha un polimorfisme extraordinari en aquest animal. Com hem esmentat més amunt, l’exemple no és únic. De l’espècie veïna Podarcis lilfordi, també de les illes Balears però localitzada a Mallorca, Menorca i illots pròxims, se n’han descrit 29 subspècies.
Altres espècies amb multitud de tàxons subspecífics a les illes mediterrànies són Podarcis sicula, principalment a Itàlia i a les illes veïnes, P. melisellensis a la costa de Dalmàcia, P. tiliguerta a Còrsega i Sardenya, P. wagleriana a Sicília, P. filfolensis a l’arxipèlag de Malta, P. peloponnesiaca a Grècia, P. erhardii a les illes de l’Egea, etc.
La Mediterrània ha estat per a la humanitat un gresol de cultures i llengües, un centre des d’on la civilització ha irradiat. Així mateix, ha estat un centre de dispersió de la fauna i, també, un gresol on han nascut noves subspècies i espècies, en aquest ambient d’elevada diversitat geogràfica i històrica.
El paper dels humans
El paisatge mediterrani es troba molt humanitzat. Els humans civilitzats han ocupat la conca mediterrània des de fa molts mil·lennis i n’han deixat l’empremta en l’entorn fins al punt que aquest paisatge no es pot explicar sense la seva acció, fins i tot en aquells casos en què aparentment no ha sofert alteracions. Aquesta intervenció ha estat duta a terme de moltes maneres diferents, de les quals potser la més antiga, persistent i eficaç ha estat l’ús del foc. Complementat amb l’agricultura itinerant o permanent, la irrigació, el pasturatge o el turisme, el foc ha deixat sentir els seus efectes en una degradació del paisatge tan antiga que actualment sembla natural. Això ha comportat la disminució o la desaparició dels boscos esclerofil·les originaris en favor de l’increment de garrigues i, en darrer terme, de formacions subestepàries com els farigolars ibèrics o el “renosterveld” sud-africà. A aquests canvis en la vegetació han seguit canvis igualment importants i menys reversibles en el sòl, que ha passat dels brunozems dels alzinars madurs a les terres rosses i rendzines de les garrigues i espinedes, fins a arribar a litosòls pràcticament nus.
La intervenció també ha tingut conseqüències sobre la fauna. Aquestes conseqüències, malgrat que són fruit d’una acció que només ha estat intensa uns quants mil·lennis, són tan importants com les que es deriven de les oscil·lacions climàtiques del Plistocè o d’altres processos que operen a escales cronològiques més àmplies. Les conseqüències indirectes es deriven de la modificació de la vegetació i d’altres aspectes de l’hàbitat originari. Les directes es deuen a la domesticació d’animals, la introducció d’algunes espècies fora de llur àrea d’origen i la persecució o eliminació d’altres. Per exemple, els grans carnívors com el lleó o el guepard eren presents a les ribes septentrionals de la Mediterrània fins a l’època de l’imperi Romà, i a les meridionals fins al començament del segle passat, malgrat que avui dia ningú no pot qualificar aquestes espècies de mediterrànies.
La domesticació dels animals (tret del gos) s’inicià als països pròxims a la Mediterrània oriental fa uns 8 000 anys. Cal destacar el fet que una de les primeres espècies domesticades és la cabra, que l’espècie humana féu servir també com a eina important en la configuració del paisatge mediterrani. La influència de la cabra, del xai i de la ramaderia en general sobre aquest paisatge ha estat, si més no, tan gran com la que les activitats humanes han tingut sobre la fauna no domesticada. Però, si bé intensa a tot arreu, aquesta influència presenta variants d’origen climàtic, topogràfic i històric. Per exemple, raons culturals han eliminat el porc de molts països de la Mediterrània i això ha privat els pagesos d’un eficaç reciclador de residus, capaç de mantenir-se en boscos adevesats que no sacrifiquen l’estructura dels alzinars originaris.
La introducció de la fauna mediterrània en altres països ja ha estat comentada més amunt. De vegades, aquesta modificació ha tingut conseqüències greus per als ecosistemes receptors i ha donat lloc a profundes modificacions en la vegetació i en la fauna locals. El cas del conill a Austràlia n’és el més conegut i evident, però les seves conseqüències, com també d’altres no tan transcendents, són força complexes i solament han estat estudiades molt superficialment. Tot i així, l’efecte principal de la intervenció humana sobre la fauna mediterrània és l’indirecte, derivat de la transformació de l’hàbitat.
La diversitat faunística ha baixat molt i ha anul·lat, en part, els efectes diferenciadors de la topografia i de les oscil·lacions climàtiques. L’incendi repetit ha eliminat els animals poc mòbils, que cerquen refugi en els arbustos, i ha afavorit els que poden fugir amb facilitat i que després tornen i s’alimenten dels brots tendres dels piròfits que apareixen després de l’incendi. Així, les tortugues terrestres dels països riberencs de la Mediterrània són abocades a l’extinció, principalment a causa dels nombrosos incendis dels seus refugis. La disminució de l’arbrat simplifica l’estructura i disminueix la varietat i l’abundor de l’entomofauna, cosa que repercuteix en les comunitats d’ocells i de mamífers insectívors. També és causa que els animals arborícoles i corticícoles cerquin una altra mena de refugis, tot fent-se lapidícoles i, en darrer terme, antropòfils i fins i tot comensals dels humans. En aquest cas, la fauna segueix les passes de la flora, moltes espècies silvestres de la qual han estat eliminades pel conreu, que ha afavorit, en canvi, l’expansió de les males herbes, invasores dels camps conreats i, en una certa mesura, també comensals preparats per a aprofitar la concentració de nutrients a la vora dels establiments humans.
El fons faunístic mediterrani
La diversitat ecològica i la fauna
Quan hom descriu l’ambient mediterrani, s’insisteix tot sovint en una de les seves característiques o en una altra, com ara l’estacionalitat, l’aridesa periòdica, els trets fisiognòmics, etc., tot seguint l’orientació del descriptor. Pel que fa als animals que l’ocupen, probablement l’aspecte que influeix més és l’elevada variabilitat de l’hàbitat en el temps i en l’espai. L’àmbit mediterrani no es caracteritza pas per la uniformitat climàtica, com és el cas de la selva equatorial, per exemple, on les diferències estacionals són mínimes; ni tampoc per l’alternança regular de les estacions activa i de repòs, com als boscos boreals de coníferes.
Generalment, els ambients mediterranis sí que presenten una certa regularitat en la successió de les tres estacions bàsiques: la curta primavera, on la natura es desvetlla del son hivernal i el paisatge es transforma en molt pocs dies, el llarg estiu-tardor, durant el qual la majoria dels animals acaben llur cicle anual o bé entren en una obligada diapausa estiuenca, i l’hivern, també curt, en què es gasten les reserves acumulades durant l’estiu i es prepara la reproducció. Malgrat tot, a aquesta regularitat general se superposen oscil·lacions, de vegades prou notables i difícils de preveure, d’humitat, temperatura i insolació. La complexitat geogràfica de les línies de costa, si més no a la conca mediterrània i a Califòrnia, el relleu, amb grans elevacions en determinats punts, i l’heterogeni mosaic de substrat rocós, sòl, vegetació i grau d’intervenció humana fan que el paisatge mediterrani sigui un dels de més difícil descripció concisa i un dels que ofereix més varietat de nínxols ecològics a la fauna.
Per tant, no és possible de descriure amb tot detall la composició de la fauna mediterrània. Les similituds entre les cinc grans àrees mediterrànies actuals (els països banyats per la Mediterrània, la part meridional de Califòrnia, la regió central de Xile, l’extrem sud d’Àfrica i la costa meridional d’Austràlia) són prou recents i sobrevingudes, i els canvis climàtics han actuat sobre bases de fauna i flora diferents. Abans de discutir amb detall el poblament animal d’aquestes àrees, caldria comparar-les a grans trets i presentar aquells elements faunístics bàsics que contenen.
Els contextos geogràfics i històrics
El sud d’Austràlia és l’àrea més peculiar; no té muntanyes, ha romàs separada durant molt de temps dels altres continents, el seu clima no és típicament mediterrani, atès que presenta una certa pluviositat a l’estiu, i la seva vegetació deriva del sotabosc dels eucaliptus i forma uns matolls (“mallee”) de composició força diferent de la resta de les àrees. La fauna que habita al “mallee” australià és majorment autòctona o bé procedeix dels deserts del nord. El paper dels humans en la configuració d’aquest paisatge fou força reduït fins a èpoques recents, però posteriorment ha estat molt intens. En aquesta intervenció, que produí l’aclariment del bosc i l’expansió dels matolls, cal incloure la introducció de fauna al·lòctona silvestre, com ara la guineu i el conill, o bé domèstica, com ara el gat i el ratolí, que, en molts indrets, esdevenen asilvestrats. Precisament, el conill prové del països de la Mediterrània, cosa que n’explica la fàcil adaptació a les condicions australianes.
La zona mediterrània de l’extrem sud d’Àfrica presenta afinitats faunístiques amb totes les altres, però sobretot amb les d’Austràlia. En aquest cas, el matoll és molt pobre i menys estructurat (“fynbos”), atès que les parts de sòl més ric han estat transformades pel conreu. Com a Austràlia, molts dels elements faunístics i florístics d’aquesta zona són d’origen tropical o subtropical, ja que, en tots dos casos, la franja adjacent de latitud més elevada (més meridional) és ocupada pel mar. El paisatge és força més heterogeni, en part a causa de la presència de cadenes muntanyoses paral·leles a la costa. Pel que fa a la fauna, sobretot la del sòl, hom pot apreciar afinitats amb els elements australians.
Les dues àrees mediterrànies americanes, a Califòrnia i Xile, són les més similars des dels punts de vista geogràfic, climàtic i ecològic. Llur situació entre la mar i una elevada serralada paral·lela al litoral, la presència de corrents freds a les aigües veïnes i la connexió recent de totes dues zones a través de l’istme del Panamà, n’ha fet força semblant el paisatge, malgrat les fonamentals diferències de fauna i flora. En tots dos casos, la fauna procedeix no solament de les zones tropicals sinó també de les zones temperades veïnes. Per últim, la influència humana també és similar en totes dues àrees, més recent que la de la conca mediterrània, però més antiga que la hom exercí a Àfrica i a Austràlia. També són força notables les diferències entre totes dues zones, sobretot pel que fa als orígens de les faunes respectives, que tenen una història biogeogràfica molt diferent.
La conca mediterrània ha donat nom a aquest tipus d’ambients. Més extensa que la resta d’àrees d’aquest tipus totes reunides, s’hi inicià el paper modificador de l’espècie humana, i d’aquí han sorgit colonitzadors i organismes que s’han establert a les altres. Des del punt de vista zoogeogràfic, constitueix un conjunt de províncies de la regió paleàrtica, amb elements faunístics derivats de les biotes temperades europees i, en un grau menor, de les biotes subtropicals africanes. La història d’aquesta regió ha estat força peculiar. La connexió entre Europa i Àfrica, ocasional per l’oest i permanent per l’est, ha permès un fàcil transvasament d’elements d’un continent a l’altre, i ha fet que la Mediterrània sigui més un nexe que no pas una barrera.
Naturalment, totes les regions de clima mediterrani tenen espècies diferents que duen a terme un paper ecològic similar. D’aquesta manera, els rosegadors excavadors del gènere Octodon a Xile es corresponen amb els dels gèneres Neotoma i Thomomys, a Califòrnia, o amb el del gènere Microtus a la conca mediterrània. Així mateix, les comunitats d’iguànids als matolls de Xile (Liolaemus) tenen llurs homòlogues a les integrades per Cnemidophorus, Sceloporus i Uta a la garriga californiana, o pels lacèrtids del gènere Podarcis a la garriga de la conca mediterrània.
El cas concret dels vertebrats
Les cinc classes de vertebrats terrestres, peixos continentals (incloent-hi àgnats, condrictis i peixos ossis), amfibis, rèptils, ocells i mamífers, es troben representades a les àrees mediterrànies, si bé els peixos del “mallee” australià són introduïts o bé procedeixen de grups marins de distribució molt general. Anant cap avall en el nivell taxonòmic, es poden trobar a les àrees esmentades uns 25 ordres de vertebrats continentals no cosmopolites; la més rica de totes és la conca mediterrània, on es donen 15 ordres. Les afinitats entre aquesta conca i l’àrea mediterrània de Sud-àfrica són fortes, ja que totes dues comparteixen nou ordres. Ben al contrari, les afinitats semblen mínimes entre Sud-àfrica i Austràlia, amb un sol ordre no cosmopolita compartit, el dels ocells fasianiformes (potser dos, si hom considera el dingo, Canis familiaris dingo, un carnívor, com a nadiu d’Austràlia, malgrat que se sap que fou introduït en temps prehistòrics). Cap dels ordres compartit no és endèmic d’una regió mediterrània. Pel que fa als ordres més àmpliament distribuïts, hi figuren els artiodàctils i els peixos cipriniformes (potser també els carnívors), que manquen sols a Austràlia, i els ocells fasianiformes, que manquen únicament al Xile central, de clima mediterrani.
Si baixem al nivell de les famílies, les pautes de distribució són força més interessants, ja que, lògicament, el percentatge de grups cosmopolites es redueix i el de grups endèmics augmenta. Representants de 179 famílies no cosmopolites de vertebrats continentals es troben en una o diverses de les regions mediterrànies. Quant als peixos continentals, la conca mediterrània i el Xile central són les regions més riques, amb set famílies, mentre que Sud-àfrica sols en té tres. Aquestes dues regions, precisament, comparteixen dues famílies, els ciprinodontiformes i els aterínids. En canvi, no hi ha cap família compartida entre Xile i Sud-àfrica. Algunes famílies són gairebé endèmiques, com ara els diplomístids i els pigídids, acantonats al Xile central, si bé ultrapassen els límits de la regió mediterrània.
Els amfibis no inclouen cap família cosmopolita (els bufònids australians són d’introducció recent) i, per tant, són particularment adients per a una comparació de faunes. Tretze famílies integren la fauna batracològica d’aquestes regions, set de les quals en la de major diversitat faunística, la conca mediterrània. En canvi, al “mallee” australià es troben sols dues famíles d’amfibis, els leptodactílids, que també són presents al Xile central, i els hílids. Entre les famílies de distribució restringida, es troben els heleofrínids, subendèmics de la regió del Cap, si bé s’estenen una mica més al nord. Totes les regions, tret d’Austràlia i Sud-àfrica, tenen famílies compartides; el nombre més gran es dóna entre la conca mediterrània i Califòrnia, que comparteixen les famílies dels bufònids, els hílids, els salamàndrids i els pletodòntids. Aquesta darrera família presenta una distribució peculiar, ja que es troba en tot l’oest dels Estats Units i al nord de Mèxic i també en àrees reduïdes d’Itàlia i de les illes tirrèniques, on ha adoptat hàbits cavernícoles. Atesa la seva escassa capacitat de migració, es tracta d’un dels millors indicadors de parentiu faunístic entre les regions mediterrànies de tots dos continents.
Els rèptils, amb una capacitat molt reduïda per a migrar i amb una dependència molt acusada de les condicions ambientals, constitueixen el grup preferit pels estudiosos biogeogràfics. Cap de les 27 famílies presents a les regions mediterrànies no és cosmopolita, si bé n’hi ha una, la dels escíncids, que té una distribució molt àmplia i disposa de representants a totes les regions, i tres famílies més, els gecònids, els colúbrids i els elàpids, que també tenen membres en quatre d’elles. Una vegada més, la conca mediterrània és la regió que en presenta una varietat més gran, amb quinze famílies, i la regió central de Xile la més petita, amb set.
Les relacions entre les regions que pertanyen al Vell Món semblen més intenses, especialment entre la conca mediterrània i Sud-àfrica, que comparteixen onze famílies de rèptils. Cadascuna d’aquestes regions comparteix, a més, sis famílies amb la zona mediterrània australiana. En canvi, la zona australiana comparteix solament dues famílies, els gecònids i els escíncids, amb Xile central, i dues també, els escíncids i els elàpids, amb la Califòrnia meridional. En aquest grup tampoc no hi ha famílies endèmiques d’una regió; això no obstant, la família dels anièlids, uns llangardaixos àpodes d’hàbits subterranis, amb només dues espècies, és subendèmica del sud de Califòrnia, i es troba a la zona de clima mediterrani i a les regions àrides properes.
Pel que fa a la facilitat de moviments, els ocells es troben a l’extrem oposat dels rèptils, de manera que és difícil de vincular-los a una àrea determinada. No és gens estrany que moltes famílies d’aus que viuen a les regions de què tractem siguin cosmopolites, i d’altres tinguin una distribució molt àmplia. Per a les 64 famílies restants, tornem a trobar una situació similar, amb les dues zones americanes una mica separades de les altres tres, que formen un conjunt coherent amb un gran nombre de famílies compartides. Dinou de les vint-i-quatre famílies que viuen a la conca mediterrània són compartides amb Sud-àfrica, que en té tretze més. En canvi, entre Austràlia i Xile, solament una família, la dels psitàcids (periquitos i cotorres), és compartida. Lògicament, cal cercar les famíles endèmiques en els ocells amb menor capacitat de moviment, és a dir, en les que no poden volar. Però no hi ha cap família d’ocells que es limiti a una sola de les regions indicades.
Quant als mamífers, la situació és una mica diferent de la registrada en els altres vertebrats. Els mamífers placentaris s’originaren en el bloc eurasiàtic a la primeria de l’era terciària, i en aquell moment Austràlia ja s’havia quedat aïllada i tenia el seu propi contingent de mamífers més antics. Per tant, per a aquest grup, les afinitats són màximes entre la Mediterrània i Sud-àfrica, com també amb Amèrica del Nord. Aquestes tres zones formen un grup, amb nexes més intensos entre les dues primeres, que comparteixen 16 de les 53 famílies de mamífers no cosmopolites ni marginals per a les regions considerades. Xile central forma un altre grup, amb només tres famílies compartides amb cadascuna de les regions anteriors. Per últim, Austràlia mostra el seu grau d’aïllament en l’escàs nombre de famílies compartides amb altres regions, cap amb Xile i només una, la dels múrids, amb la Mediterrània i Sud-àfrica. Aquestes últimes zones contenen més de vint famílies no cosmopolites cadascuna, mentre que Austràlia, per les raons apuntades més amunt, només en té set.
L’anàlisi podria continuar i es faria cada vegada més complexa, si hom baixés al nivell dels gèneres o al de categories taxonòmiques inferiors. En aquest cas, minvaria el nombre de grups cosmopolites, fins i tot entre els ocells, i augmentaria el nivell d’endemisme. En efecte, hi ha força gèneres i moltes espècies la distribució dels quals se circumscriu a una sola regió mediterrània. Els casos més notables es troben entre els amfibis i els rèptils, a causa de llur dependència del medi. Espècies com ara el ferreret (Alytes muletensis), d’àrea extremament reduïda, o l’espècie israeliana Discoglossus nigriventer, de la mateixa família (si és que encara n’hi ha, ja que fa dècades que no se n’ha vist cap exemplar) són exemples d’endemismes mediterranis extrems. Entre els gèneres, hom pot esmentar Pleurodeles o Speleomantes, dos urodels limitats a la conca mediterrània occidental. Altres gèneres il·lustren els lligams biogeogràfics entre les diferents regions: així, els gèneres Hystrix (porcs espins) i Genetta (genetes), presents a la Mediterrània i a Sud-àfrica, són indicadors addicionals de l’afinitat entre totes dues regions.
La inclusió dels invertebrats en aquesta anàlisi permetria, lògicament, d’enfortir-ne les conclusions. La fauna del sòl ha estat analitzada amb aquests criteris i hom ha posat de manifest els diferents graus d’afinitat entre les cinc regions, amb resultats similars als obtinguts en el cas dels vertebrats: la importància de la fauna compartida és màxima entre la Mediterrània i la regió del Cap, i també amb el sud de Califòrnia. Xile central presenta una certa afinitat amb aquesta darrera regió, i el sud d’Austràlia ofereix la seva peculiaritat en totes les anàlisis. La concordança reflecteix la història dels moviments continentals i les relacions canviants d’accessibilitat de les diferents regions.
Les afinitats zoogeogràfiques mediterrànies
Les regions de clima mediterrani comparteixen no solament un clima comú, sinó també una fisiognomia de vegetació, paisatge i ús del sòl semblants. Aquest fet és notable, si hom té en compte que les cinc regions mediterrànies del món estan distribuïdes d’una manera disjunta i separades per grans distàncies, i que, per això, han desenvolupat les seves pròpies biotes en aïllament i a partir d’estocs filogenètics fonamentalment diferents. Tanmateix, no hi ha cap altra disjunció terrestre mundial que presenti tantes similituds com la formada per les regions mediterrànies.
Des del punt de vista zoològic, i igual que passa amb la flora, les similituds no són degudes principalment a la fauna compartida, atès que no hi ha espècies animals comunes a les diferents regions, si n’exceptuem algunes, poques —bàsicament d’invertebrats—, que han aconseguit dispersar-se a llarga distància, sovint amb la intervenció dels humans (tot i així, solen tenir una àmplia distribució, són cosmopolites o, fins i tot, ubiqües, i no característiques de les àrees mediterrànies). En les categories taxonòmiques superiors (gènere, família, etc.), la similitud faunística (és a dir, el nombre de tàxons compartits) entre dues o més regions s’incrementa gradualment, sobretot al nivell jeràrquic de família. A més, segons el nivell considerat, les afinitats poden variar. Així, per exemple, Califòrnia i Xile són les regions que més s’assemblen quant al nombre de gèneres d’ocells compartits, mentre que Sud-àfrica no en té cap en comú amb cap d’aquestes; en canvi, al nivell de família, les més pròximes són Califòrnia i la Mediterrània, i les més diferents són Xile i la Mediterrània.
D’altra banda, la similitud entre les cinc regions queda més clarament reflectida en certes manifestacions de convergència ecològica i morfològica de les espècies i les comunitats animals.
Els grups faunístics de la conca mediterrània

Tallarols i tallaretes són ocells del gènere Sylvia, de mida petita i, en general, de colors apagats. La meitat de les espècies d’aquest gènere són exclusivament mediter- rànies, representen un dels pocs grups d’ocells autòctons i probablement el cas més notable de radiació adaptativa d’ocells en aquesta regió, sobretot a la part occidental, on presenten la major diversitat. Aquesta espe- ciació degué ser afavorida per la fragmentació de la vegetació i els consegüents aïllaments geogràfics al llarg dels períodes glacials i interglacials del Plistocè. Al dibuix, tallareta vulgar (Sylvia communis), espècie no exclusivament mediterrània, ja que s’estén per tot Europa i arriba fins a Sibèria occidental, tallarol trencamates (Syl- via conspicillata), tallarol de garriga (Sylvia cantillans), tallareta sarda (Sylvia sarda), tallareta cuallarga (Sylvia undata), tallarol emmascarat (Sylvia hortensis) i tallarol capnegre (Sylvia melanoce- phala).
Marisa Bendala / ECSA
Resulta difícil definir què és una espècie característica de la regió mediterrània, atesa la seva situació de cruïlla biogeogràfica, especialment acusada durant els períodes glacials i interglacials, en què hi hagué importants desplaçaments de vegetació i successives invasions de fauna. Una espècie de la Mediterrània pot tenir històries molt diferents. Pot haver evolucionat sota un clima mediterrani o bé abans de l’aparició d’aquest clima, però s’hi ha pogut pre-adaptar. També pot tractar-se d’una espècie immigrant que hagi colonitzat la regió defugint condicions molt àrides o molt fredes; finalment, pot ser una invasora que ha arribat recentment. Per tant, la Mediterrània és un mosaic de formes de diferents orígens biogeogràfics.
L’àrea mediterrània presenta una gran riquesa d’ocells. Entre els ocells nidificants terrestres domina un element format per espècies que ultrapassen els límits de la formació esclerofil·la i s’endinsen en els boscos caducifolis de la regió paleàrtica. Un altre element s’estén per les praderies d’Euràsia i les estepes del sud i el sud-est. Finalment, els endemismes mediterranis només representen un conjunt discret. D’altra banda, un seguit de grups tropicals, com ara els cràcids, els psitàcids, els trogònids i d’altres, que poblaven Euràsia durant el Miocè (als quals s’afegiren els vultúrids en el Pliocè) es van extingir totalment a la Mediterrània abans del Plistocè, raó per la qual són rares les relacions faunístiques de la regió amb àrees exteriors a Euràsia i nord d’Àfrica. Durant el Plistocè es van acumular en repetides ocasions a la Mediterrània tots els tipus faunístics d’Europa, i cada vegada que retrocedien les glaceres, expandien les seves àrees cap al nord, però sense abandonar la conca mediterrània, de manera que aquesta quedava sempre connectada faunísticament amb l’Europa central, sense que s’arribés a produir un aïllament, ni tampoc especiacions importants. Per aquest motiu, hom considera que l’ornitofauna mediterrània és essencialment una part de l’ornitofauna paleàrtica, però empobrida a causa de la seva situació peninsular respecte a Euràsia, i amb relativament pocs endemismes. De fet, la conca mediterrània en l’actualitat és un quarter d’hivern per a una bona part de l’avifauna eurasiàtica.
Pel que fa als mamífers, hom troba no menys de cent-cinquanta espècies mediterrànies, a més de mig centenar d’espècies que tenen una àrea de distribució reduïda a la regió. L’intercanvi de faunes entre Europa i Àfrica ha estat i continua essent molt poc freqüent. Durant la major part del Quaternari els mamífers de la riba sud evolucionaren independentment dels de la riba nord. Anteriorment, tot i que les dues ribes estaven connectades per ponts terrestres durant la crisi messiniana, les dades paleontològiques indiquen que només set espècies de rosegadors i un lagomorf migraren d’Europa a l’Àfrica a través d’Espanya (alguns d’ells sobrevisqueren fins al final del Pliocè), mentre que cinc gèneres de rosegadors africans colonitzaren amb èxit la Península Ibèrica. Més tard, durant el Pliocè i el Plistocè, l’estret de Gibraltar impedí intercanvis posteriors. Tot això fa que la major part de la fauna del sud d’Europa sigui d’afinitat eurosiberiana i mediterrània, mentre que la major part de la nord-africana és etiòpica, és a dir, d’origen africà sudsaharià. Els elements secundaris de la mastofauna mediterrània serien l’iranoturanià (comprèn els deserts freds d’Àsia central), el saharosindià (un cinturó de deserts que va des del Marroc fins al Sind, a l’Índia), i en últim lloc, l’oriental, format per Àsia al sud de l’Himàlaia. Finalment, els autèntics endemismes mediterranis es diferenciaren essencialment a partir de faunes insulars empobrides, durant centenars de milers d’anys d’aïllament. El porc espí d’Itàlia (Hystrix cristata), és l’últim arcaisme europeu d’origen tropical, però no difereix de les poblacions africanes; la resta de les espècies arcaiques s’extingiren abans de completar la seva diferenciació en endemismes.
Els grups faunístics de les altres regions mediterrànies
De les cinc regions del bioma, la conca mediterrània és la que ha conegut un grau més baix d’aïllament biogeogràfic i evolutiu, seguida de Califòrnia, Xile (tancada entre el desert d’Atacama i els Andes), Sud-àfrica i, finalment, Austràlia.
Una part dels ocells de la Califòrnia mediterrània està confinada al “chaparral”, però una proporció encara més gran té afinitats amb la zona boreal d’Amèrica del Nord, mentre que gairebé tota la resta presenta una àmplia distribució al sud-oest dels Estats Units i el nord-oest de Mèxic. A Xile, pràcticament no hi ha ocells propis del matorral, sinó que s’estenen al bosc temperat de Nothofagus del sud i als Andes, o bé tenen una àmplia distribució a Sud-amèrica o al Nou Món; per tant, l’ornitofauna xilena es degué originar a parts iguals en els Andes tropicals i en el bosc temperat del sud; a aquest model s’afegeixen algunes espècies cosmopolites.
La majoria d’espècies d’ocells del “fynbos” sud-africà són compartides al nord i a l’est amb la regió del Karoo, de vegetació semidesèrtica, mentre que d’altres ho són amb les praderies d’Orange i de Bostwana, i algunes altres amb les àrees relictes de bosc a la costa.
Pel que fa als llangardaixos, Xile comparteix les seves dues úniques famílies amb Califòrnia, la qual en té dues més de pròpies. Per contra, la Mediterrània no comparteix cap família amb les dues anteriors.
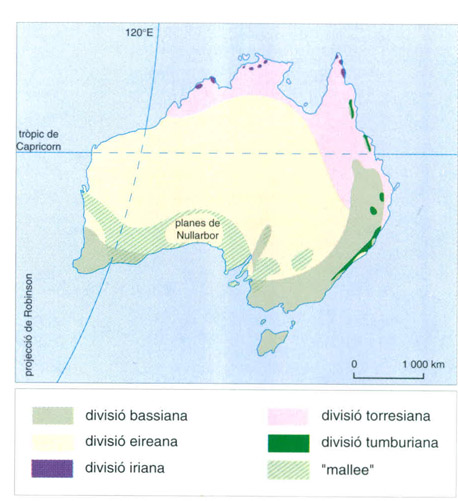
Distribució de les comunitats d’ocells del “mallee” en relació amb les divisions zoogeogràfiques de l’avifauna australiana. La divisió coincideix en part amb la dels tipus de vegetació d’Austràlia.
Editrònica, a partir de fonts diverses
Les afinitats de la fauna de vertebrats del “mallee” australià són intracontinentals, a conseqüència del seu aïllament més marcat. Tan sols quatre ocells passeriformes i un llangardaix, altament especialitzats, hi estan confinats, és a dir, en són propis i característics. En comparació amb la fauna de les zones veïnes de bosc i bruguera, el “mallee” presenta una diversitat de llangardaixos més gran, però, en canvi, mostra una diversitat inferior d’ocells i amfibis, mentre que és similar la dels mamífers. Les granotes i els gripaus tenen una escassa representació en el “mallee”, i no hi ha cap endemisme; a més, no hi ha espècies comunes a les dues àrees que formen la regió separades per les planes desèrtiques del Nullarbor, que actuen de barrera a la dispersió, tot i que alguns gèneres originaris de l’àrea sud-oriental aconseguiren envair el sud-oest durant els períodes interglacials. Pràcticament totes les famílies de rèptils del continent australià tenen representants al “mallee”, la majoria dels quals penetren en la zona àrida anomenada d’Eyre, i la resta ho fa en la zona dita de Bass, més humida (sud-oest i sud-est d’Austràlia, a més de Tasmània). De les 163 espècies d’ocells que habiten el “mallee” d’una manera permanent o estacional, algunes en són endèmiques, però la gran majoria pertanyen a aquestes dues regions (llac Eyre i estret de Bass), excepte unes sis, que es relacionen amb la zona de Torres, que representa el nord i el nord-est del continent. Finalment, set famílies de mamífers, amb prop de 30 espècies, són característiques de l’Austràlia mediterrània, però cap d’elles no hi està especialment adaptada; per la seva banda, el reduït nombre d’espècies es deu, si més no en part, a les extincions causades pels humans en el decurs dels darrers segles.
El cas de la fauna edàfica mediterrània
Graus d’afinitat entre els pseudo-escorpius, a nivell de família i gènere, reflectits en el gruix de les sagetes. Si bé no hi ha una fauna compartida a les regions amb clima mediterrani, es poden trobar algunes afinitats entre elles. Aquest antic grup assoleix les seves majors diversitats precisament en els sòls i les cavernes de regions mediterrànies. Hom pot observar la importància més gran de les relacions “horitzontals” respecte a les “verticals”, com a testimoni d’un llegat geològic comú molt antic en què les formes arcaiques boreals evolucionaren aïllades de les australs sobre els continents de Lauràsia i Gondwana. En contrast, les afinitats verticals són més recents i majors entre Califòrnia i Xile que entre la Mediterrània i Sud-àfrica, ja que el Sàhara ha estat una barrera forta per a les faunes dels sòls humits mentre que al Nou Món els Andes han pogut servir de pont per a certes migracions latitudinals. Finalment, no hi ha espècies comunes entre les regions, excepte algunes de cosmopolites, probablement introduïdes pels humans. Aquest patró d’afinitats es correspon bastant bé amb el d’alguns altres grups d’invertebrats del sòl, però no amb els de la majoria de vertebrats.
Editrònica, a partir de Di Castri, 1981
La fauna del sòl és particularment interessant i diversa en els sòls mediterranis. Alguns grups, com ara el dels pseudoescorpins, es troben a les cinco regions, i hi tenen una diversitat comparable a la de la pluviïsilva tropical. En canvi, d’altres són secundaris, com és el cas dels tèrmits, la importància dels quals va disminuint des de Sud-àfrica i Austràlia, a Califòrnia, la conca mediterrània i Xile, paral·lelament al grau d’aïllament de cada regió respecte a les influències del clima plujós tropical.
Artròpodes higròfils dels grups més diversos han trobat en la vida en el sòl una solució per a evitar les contrastades condicions extremes diàries i estacionals del clima, atès que la seva limitada capacitat de dispersió no els permet migrar estacionalment cap a regions més humides. Aquesta estratègia adaptativa es dóna en grups arcaics de col·lèmbols i proturs, i ha estat adquirida en grups més moderns de coleòpters, de pseudo-escorpins, i d’altres, que evolucionaren secundàriament a partir d’ancestres epigeus en el sentit d’una reducció de la talla, l’aparició de l’apterisme, l’anoftàlmia i altres adaptacions a la vida edàfica. Tanmateix, les principals adaptacions no són específiques dels ambients edàfics mediterranis, sinó que ja existien en la fauna del sòl abans de l’aparició del clima mediterrani, i actualment es poden trobar fora d’aquestes regions.
Les afinitats filogenètiques entre les faunes del sòl de les diferents regions mediterrànies són importants. Com a resultat dels canvis geològics i paleoclimàtics, s’han anat acumulant línies filogenètiques de diversa procedència. Com a il·lustració d’aquest origen híbrid, hom pot considerar el cas dels sòls xilens, en els quals destaca un element biogeogràfic paleoantàrtic o austral (que comprèn Sud-amèrica, Austràlia, Nova Zelanda, Sud-àfrica i les illes antàrtiques), que és present en nombrosos grups: els gammàrids, els col·lèmbols, els coleòpters, els turbel·laris, els oligoquets, els isòpodes, els pseudo-escorpins, els procòpters i d’altres. Un segon grup, format bàsicament per espècies xeròfiles epigees d’hàbitats àrids com ara solífugs, aranyes, tenebriònids, pseudo-escorpins, alguns formícids i àcars trombidiformes, està molt relacionat amb distribucions andines, i forma l’element neotròpic, probablement originat per línies neotròpiques i línies àrtiques que entraren a Xile per la serralada procedents del nord. L’element pantropical, és a dir, distribuït en les regions tropicals de la Terra, és rar en els sòls xilens, i només representat per alguns pseudo-escorpins, queiridioidèids, alguns diplòpodes i diversos nemàtodes paràsits. La major part de les línies arcaiques degueren tenir una distribució aproximadament contínua en el Mesozoic i actualment formen una disjunció mundial molt fragmentada: es tracta de relíquies biogeogràfiques, la majoria estrictament higròfiles i, per tant, limitades a les capes profundes dels sòls mediterranis; aquest seria el cas de gran part dels proturs, pauròpodes, sínfils i palpígrads. Finalment, un últim element és el cosmopolita, compost d’una banda per espècies amb bona capacitat de dispersió passiva —facilitada per tenir de formes de resistència i una mida petita (rotífers, tardígrads i molts protozous)— i, d’altra banda, per espècies introduïdes accidentalment per l’espècie humana en l’expansió de les seves pràctiques agrícoles, amb una gran capacitat de colonització, i que són el component principal dels sòls conreats, sobretot els de regadiu, com ara acàrids, col·lèmbols, oribàtids, nematodes, lumbrícids europeus i algun isòpode.
Les introduccions i les invasions
El fet que tota àrea de distribució sigui dinàmica fa que hom consideri la invasió no com un esdeveniment, sinó com un procés allargat en el temps. Les espècies potencialment invasores solen ser generalistes, és a dir, solen presentar una tolerància ecològica més gran. Tot i que és un procés normal en la natura que les àrees de distribució dels tàxons s’expandeixin, aquest procés s’ha vist accelerat o forçat en temps històrics per les possibilitats de dispersió dels organismes a llarga distància facilitada pels humans. Alguns autors consideren que els ecosistemes i les regions que en temps geològics i històrics han sofert períodes llargs de pertorbacions climàtiques, humanes, etc., són més resistents a la invasió de formes al·lòctones, i alhora poden tenir més espècies capaces d’envair altres regions; la regió mediterrània en seria un exemple.
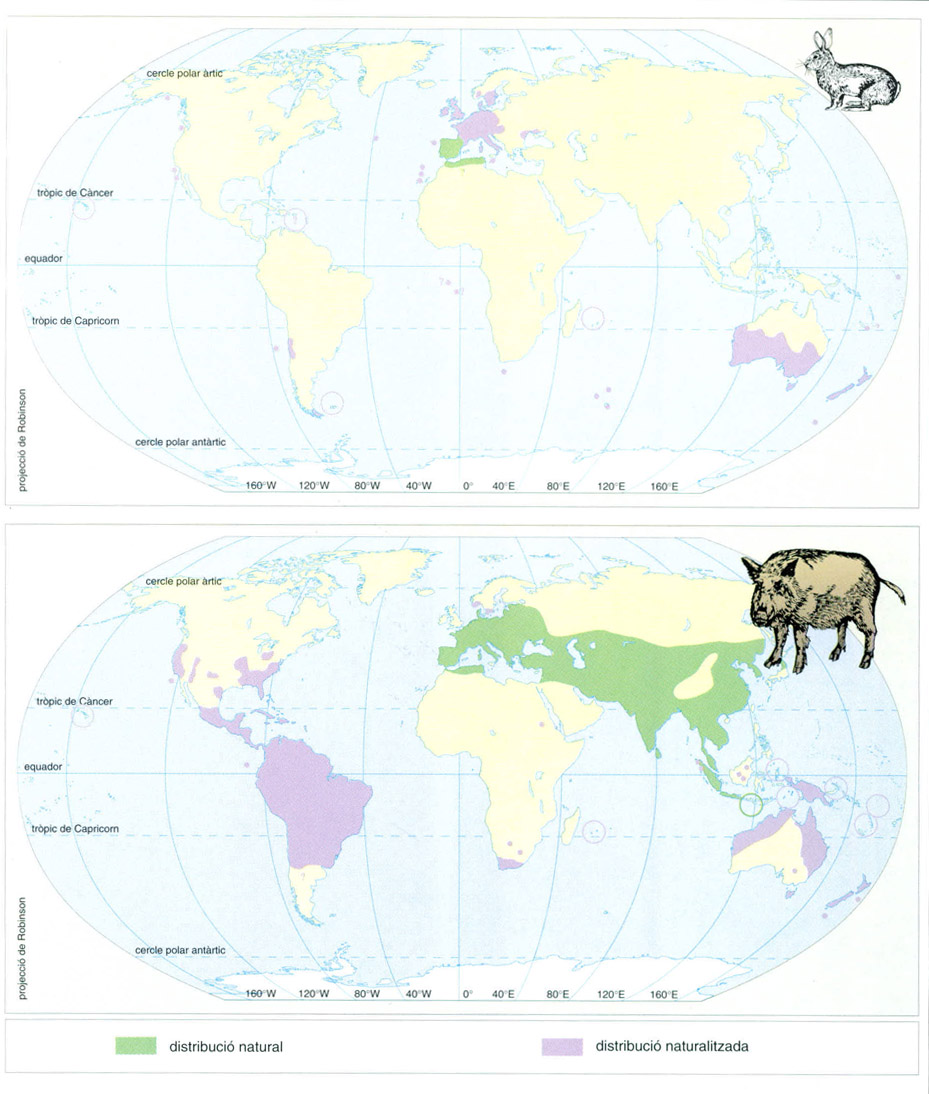
Àrea de distribució actual al món del conill (Oryctolagus cuniculus) i del porc senglar (Sus scrofa), indicant-hi la seva àrea natural i la que ha estat forçada pels humans. En la seva àrea naturalitzada els animals viuen i es reprodueixen en el nou territori com si fos la seva àrea natural.
Editrònica, a partir de “Naturalized mammals of the world”, Lever, 1985
Els humans han introduït d’una manera intencionada o fortuïta un seguit d’espècies generalistes d’àmplia distribució, algunes de les quals són cosmopolites ubiqües, en ecosistemes originàriament formats per espècies amb una distribució i uns requeriments més restringits. Pel que fa als mamífers, a Austràlia meridional s’ha intentat introduir durant els darrers dos segles, amb èxit desigual, una cinquantena d’espècies. A Califòrnia, algunes introduccions han assolit una distribució molt limitada en el “chaparral”, i altres són comensals de l’espècie humana o habiten àrees suburbanes, tot i que mostren una certa expansió silvestre. Algunes espècies domèstiques també han tornat al medi silvestre: és el cas del cavall (Equus caballus), l’ase (E. asinus), la cabra (Capra hircus), el bou (Bos taurus), el be (Ovis aries), l’esquirol de les Rocalloses (Sciurus carolinensis), el gat (Felis catus) i el gos (Canis familiaris). A Sud-àfrica, l’impacte assolit per les plantes introduïdes contrasta amb l’escassa rellevància de les introduccions de mamífers. Així, la rata negra (Rattus rattus), la rata comuna (R. norvegicus), el ratolí domèstic (Mus musculus) i l’esquirol (Sciurus carolinensis) són tots comensals, és a dir, viuen només en proximitat dels humans i sense expansió en el “fynbos”; el conill (Oryctolagus cuniculus) únicament ha estat introduït en illes que no tenen vegetació de “fynbos”, mentre que ni el senglar (Sus scrofa), ni la daina (Dama dama) no han esdevingut asilvestrats; tan sols el tahr (Hemitragus jemlahicus) ha estat un mamífer realment invasor, però sempre d’una manera molt local.
La majoria de les espècies d’ocells invasors són comensals de l’espècie humana i de les seves activitats en totes les regions mediterrànies. Hom calcula que menys del 5% de l’avifauna mediterrània de Califòrnia, Xile, Sud-àfrica i Austràlia és potencialment invasora. A diferència d’altres grups, com per exemple les plantes, els mamífers, els insectes paràsits i altres organismes causants de malalties, la dispersió deguda als humans no ha estat un factor important en les invasions d’ocells a la Mediterrània. De fet, molt poques han tingut èxit després d’arribar, i encara són menys les que han assolit una rellevància ecològica dintre de la fauna mediterrània; per contra, entre les espècies natives, la major part han sofert canvis en les seves àrees a conseqüència de les modificacions del paisatge per l’acció humana.
El caràcter insular de l’avifauna xilena, aïllada pels Andes dels intercanvis amb altres faunes, fa que el seu contingent d’espècies sigui clarament inferior al de Califòrnia. De les introduccions practicades en la primera, dues espècies es poden considerar ben establertes i amb un impacte ecològic sobre l’avifauna; en la segona, tan sols l’estornell (Sturnus vulgaris) mereix aquesta consideració. Finalment, tres de les introduccions ben establertes són comunes a ambdues regions, l’esplugabous (Bubulcus ibis), el colom (Columba livia) i el pardal comú (Passer domesticus), però totes tres es troben àmpliament distribuïdes en el món i, per tant, s’han de veure com a cosmopolites ubiqües i no com a pròpies d’àrees mediterrànies.
A Sud-àfrica crida l’atenció l’elevat nombre d’introduccions que han fracassat. Cap espècie nativa d’un altre continent no és present d’una manera regular en el “fynbos” no degradat, mentre que en canvi sí que n’hi ha casos en zones de “fynbos” alterades pels humans; únicament podrien tenir un cert impacte ecològic en el “fynbos” un parell d’espècies africanes i l’estornell (Sturnus vulgaris), procedent d’Europa. Per acabar, al “mallee” australià, del centenar d’introduccions que hom hi ha fet, només una dotzena estan ben establertes, i gairebé totes són espècies generalistes en les seves àrees d’origen, d’àmplia distribució a Europa o Àsia, i filogenèticament modernes.
Els fenòmens de convergència
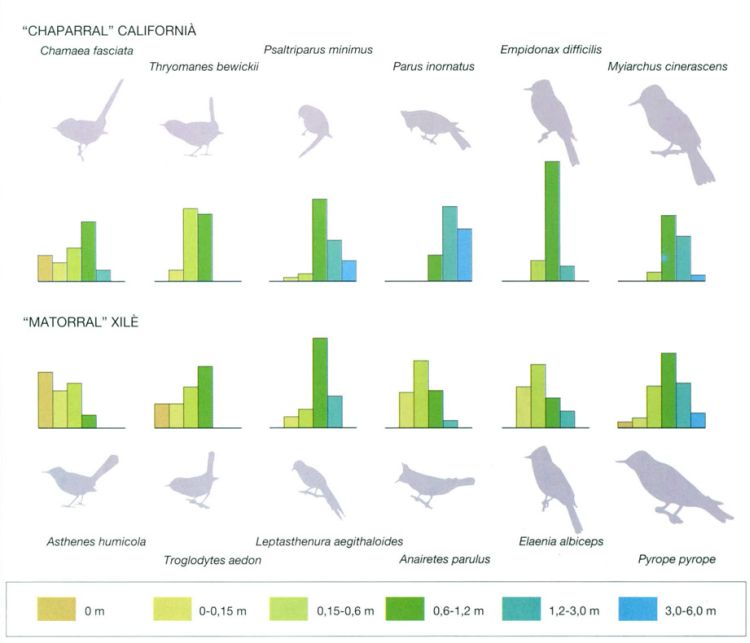
Convergència ecològica en parelles d’espècies d’ocells insectívors de les formacions esclerofil·les de Califòrnia i de Xile. Els diagrames de barres indiquen les freqüències de distribució de cada espècie a les diferents alçades (en metres) on troben aliment. Hom pot observar que espècies de gèneres i famílies diferents són equivalents ecològics no sols per la similitud de l’estrat de vegetació on s’alimenten, sinó també per la categoria tròfica a la qual pertanyen (les quatre primeres parelles són insectívores del fullatge i les altres dues cacen insectes al vol), com també per les mides semblants (les siluetes són totes a la mateixa escala).
Jordi Corbera, a partir de fonts diverses
Quan organismes d’un mateix tàxon queden aïllats genèticament i geogràficament, tenen tendència a diferenciar-se progressivament en un o més caràcters a mesura que evolucionen independentment. Tanmateix, no tots els caràcters que es modifiquen en els tàxons aïllats divergeixen necessàriament; per això, espècies no emparentades que viuen en àrees separades geogràficament però amb característiques ambientals similars, poden desenvolupar, d’una manera independent, morfologies i estratègies ecològiques semblants per a adaptar-se a uns medis també semblants. Aquest tipus d’evolució és el que hom anomena convergència o evolució paral·lela (els tàxons que la presenten reben el nom d’equivalents ecològics), i és aplicable no solament al fenotip (aspecte morfològic, de comportament, etc.) de les espècies, sinó també a d’altres de més subtils, com ara la distribució i la densitat de les poblacions, la diversitat, la utilització dels recursos o l’organització d’una comunitat.
Quan hom compara dins d’una mateixa categoria tròfica el nombre d’espècies i la densitat d’individus per hectàrea entre l’avifauna de diferents àrees mediterrànies, hom observa similituds en l’organització de la comunitat. Per exemple, dues regions poden diferir clarament pel nombre d’espècies dins d’una mateixa categoria tròfica, com ara la categoria dels ocells que s’alimenten a terra, però, en canvi, reduir la diferència existent en la densitat total d’individus de la mateixa categoria, gràcies al fet que la regió més rica d’espècies té una mitjana de densitat per espècie inferior a la més pobra; així, l’excés d’espècies d’una categoria queda parcialment compensat per una densitat específica menor.
En general, i també en altres grups, com ara els llangardaixos, la convergència es manifesta més a l’escala de les comunitats que no pas a la de les espècies, atès que una espècie no té necessàriament un equivalent morfològic i ecològic en altres regions, mentre que sí que hi pot haver una distribució semblant a l’escala dels grups tròfics i dels comportaments en la recerca de l’aliment, encara que amb les particularitats pròpies de cada regió; per exemple, sembla que els ocells de Califòrnia es divideixen la regió per hàbitats, mentre que els xilens ho fan per àrees geogràfiques, a causa de l’aïllament molt més gran del país.
Totes les adaptacions convergents esmentades són recents, i en qualsevol cas, posteriors a l’aparició en el món del clima mediterrani. Tanmateix, la convergència i les seves causes encara no es coneixen gaire bé. Hi ha, d’altra banda, certes discrepàncies, és a dir, casos en què, per exemple, l’espai morfomètric que descriu l’avifauna d’una regió no mediterrània presa com a control es pot sobreposar completament a l’espai corresponent de les regions mediterrànies, fet que no pot ser interpretat com un fenomen de convergència entre faunes, ja que provenen de medis amb clima i vegetació diferents.
Finalment, hi ha uns determinats tipus ecològics que no estan representats en totes les regions mediterrànies per raons històriques que transcendeixen els límits de l’adaptació ecològica de les espècies. Per exemple, la conca mediterrània és l’única de les cinc regions del bioma que no té ocells que s’alimentin del nèctar de les flors; a la regió Paleàrtica les flors no estan especialitzades a ser pollinitzades per ocells que mengen nèctar, sinó a ser pol·linitzades per insectes. Aquesta antiga coevolució entre insectes i plantes, juntament amb les restriccions imposades pel programa de desenvolupament d’un animal (els ocells en aquest cas), fan que les flors de la conca mediterrània quedin fora dels límits de plasticitat adaptativa dels nostres ocells.
Més espai per als petits que no pas per als grossos
Un mosaic de nínxols ecològics
Hom ja ha destacat la complexitat del bioma mediterrani davant la uniformitat d’altres biomes, com ara la taigà o els deserts. Aquesta complexitat es desenvolupa en diversos nivells, des de les grans unitats geogràfiques fins als petits micronínxols determinables tan sols després d’una anàlisi elaborada de l’estructura dels ecosistemes. La divisió principal és de caire geogràfic i permet de diferenciar les cinc regions de clima mediterrani prou conegudes, amb faunes d’origen biogeogràfic diferent i de composició diversa. Algunes d’aquestes regions se subdivideixen en àrees relativament independents i amb trets més homogenis, com és el cas d’Austràlia, amb dues àrees mediterrànies separades pel desert de Nullarbor, o en el cas de les àrees ibèrica, itàlica, balcànica, pòntica, anatòlica i africana, a la conca mediterrània. Sota aquest nivell, les divisions principals són de caire climàtic i depenen de la latitud i de l’altitud. Per exemple, hom pot distingir comunitats xerotermodinàmiques, termomediterrànies, mesomediterrànies o submediterrànies, segons la durada dels respectius períodes àrids.
La subdivisió pot prosseguir d’acord amb criteris principalment topogràfics: les grans serralades, les conques dels grans rius, les illes principals, constitueixen classes d’unitats amb composicions faunístiques més o menys diferents. A llur torn, aquestes unitats se subdivideixen en àrees menors, una vall, una serra, fins i tot, en algun cas, un vessant o un cim. Per dessota d’aquest nivell, hi ha els elements del paisatge, com ara un bosc concret, però la subdivisió pot continuar fins a arribar a espais de dimensions mil·limètriques o encara més petits. Aquest és el patró general de la complexitat ecològica, constituïda partint d’estructures dins unes estructures més grans, com les nines russes encaixades l’una dins l’altra, però amb la complicació addicional que els nivells no es troben separats, sinó que les fronteres són difuses i en un mateix nivell s’inclouen unitats d’entitat diferent.
Alguns nínxols exemplars
El recurs més utilitzat en la caracterització de nínxols ecològics és l’alimentació, i les altres dimensions del nínxol acostumen a subordinar-s’hi. Quan hom analitza l’estructura d’un ecosistema, els animals que l’integren solen separar-se d’acord amb llur forma d’alimentació, i també segons el lloc i el temps en què la duen a terme. Hom sobreentén que els animals herbívors no competeixen per l’aliment amb els carnívors, que els que s’alimenten a les capçades dels arbres no competeixen amb els que ho fan a terra, i que els que mengen de nit no interfereixen amb els que mengen de dia. D’aquesta manera, es defineixen nínxols que, d’acord amb el principi d’exclusió, acostumen a estar ocupats per una única espècie. A més, hom accepta que dos animals morfològicament molt diferents, com ara un ocell i un amfibi, no competiran encara que tinguin hàbits alimentaris paral·lels, ja que posseeixen uns mitjans molt diferents per tal d’aconseguir menjar. Així, hom pot parlar de nínxols ecològics per als ocells, els mamífers, etc., que hom pot designar amb un mateix terme.
Vegem alguns exemples de nínxols ecològics ocupats per animals semblants als biomes mediterranis. Naturalment, es tracta només d’una relació sumària, amb una insistència especial en els ocells, grup que ha estat força més estudiat que la resta. Podem començar amb l’exemple que va servir el 1915 per a la definició del terme “nínxol ecològic”. Es tracta d’un ocell, Toxostoma redivivum, que cerca el seu aliment al sòl, sota una coberta vegetal densa, omnívor, amb plomatge adequat per a passar desapercebut en aquest entorn, que, si bé es pot trobar en diverses associacions vegetals, predomina en els matolls de pota de cavall de Califòrnia. Als matolls xilens, una espècie de mímid, Mimus thenca, presenta les mateixes característiques i ocupa el mateix nínxol. El nínxol homòleg a la conca mediterrània és ocupat per la merla (Turdus merula). A Austràlia, Serinus cautus ocupa un nínxol similar, però no idèntic, i l’espècie corresponent a Sud-àfrica és el túrdid Cossypha caffra.

Caprimúlgids típics de cada un dels quatre àmbits mediterranis més conspicus: l’enganyapastors (Caprimulgus europaeus) de la conca mediterrània, Phalaenoptilus nuttallii de la mediterrània californiana, C. longirostris del Xile central i C. pectoralis de la regió capenca. Cap d’aquestes espècies no és exclusiva de l’àrea mediterrània on es troba, però cadascuna hi fa el mateix paper ecològic depredador pel que fa al vol d’insectes crepusculars.
Lluís Sanz
Altres ocells s’agrupen en gremis que ocupen nínxols afins. Per exemple, hom pot definir un nínxol per als ocells insectívors que s’alimenten entre el fullam, un altre per als nectarívors, un altre per als que cerquen l’aliment als troncs i a les escorces, etc. El resultat més interessant d’aquesta distribució és que cada nínxol o grup de nínxols té aproximadament el mateix nombre d’espècies i la mateixa densitat d’individus en regions molt allunyades. Així, hi ha un ocell insectívor d’hàbits crepusculars, que caça les preses al vol, que pertany a la mateixa família dels caprimulgiformes, tant a la conca mediterrània (l’enganyapastors, Caprimulgus europaeus) com a Califòrnia (Phalaenoptilus nuttallii), a Xile central (Caprimulgus longirostris) o a Sud-àfrica (Caprimulgus pectoralis). De la mateixa manera, el grup dels que s’alimenten de fruits i llavors consta de dues a quatre espècies, segons les regions, però en tots els casos es tracta d’ocells de les mateixes dimensions, amb una densitat semblant a tot arreu, aproximadament un individu per hectàrea.
Evidentment, els nínxols s’estableixen també per als mamífers, que no competeixen amb els ocells perquè llur activitat és predominantment nocturna. Hi ha nínxols homòlegs per a mamífers a les cinc regions de clima mediterrani: així, els ratolins de bosc són representats a Europa per Apodemus flavicollis, a Amèrica del Nord per Peromyscus californicus, a Sud-àfrica per Myomiscus veroxii, a Xile per Akodon longipilis i a Austràlia per Pseudomys albocinereus. Unes llistes semblants es podrien establir per a insectívors cavadors, per a petits carnívors, per a petits rat-penats insectívors, etc. En cada cas, el nínxol és parcialment compartit per altres espècies afins. La similitud entre determinats nínxols de les regions mediterrànies es posa de relleu atès l’èxit d’algunes introduccions; és el cas, per exemple, de la daina (Dama dama), un cèrvid originari del sud d’Europa, que s’ha introduït tant a Califòrnia com a Xile, Sud-àfrica i Austràlia i que, a tot arreu, s’ha integrat en els ecosistemes locals, duent a terme el mateix paper que a la seva zona d’origen.
Els rèptils formen un grup molt adient per a l’anàlisi de la distribució de l’hàbitat en nínxols separats i, de fet, han servit per als primers estudis d’aquesta mena, duts a terme al subdesert i al “mallee” australians i que, posteriorment, s’han estès a altres àrees similars arreu del món. Si bé les famílies dominants de petits llangardaixos són diferents d’unes regions a unes altres, les analogies es poden trobar en totes les àrees de clima mediterrani. Les petites sargantanes europees s’inclouen a la família dels lacèrtids, mentre que les d’Amèrica pertanyen a les famílies dels tègids i dels iguànids, les de Sud-àfrica són escíncids i les d’Austràlia, principalment agàmids. Però a tot arreu es tracta d’animals que viuen al sòl, que es mouen ràpidament, que romanen actius sols durant les hores de calor i que s’alimenten gairebé exclusivament d’insectes. En molts indrets, conviuen dues espècies pertanyents al mateix gènere, que es reparteixen els recursos eficaçment i que molt poques vegades es barregen. Els temes apreciats pels ecòlegs actuals, com ara convergència i substitució de faunes, compensació de densitat, estructura d’hàbitat, encobriment de nínxols, etc., han estat investigats preferentment en els llangardaixos.
Atesa la gran varietat d’espècies d’insectes que hi ha al bioma mediterrani, no és estrany que es tracti d’un grup que ha desenvolupat graus extrems d’especialització en un nínxol estricte. Hi ha insectes que viuen sense cap problema en aigua salabrosa o fins i tot salada, que viuen a la cavitat nasal dels amfibis o bé que entren i surten per l’orifici respiratori dels llimacs, que passen tota llur vida sobre la flor on han nascut, o que s’alimenten exclusivament dels dipòsits d’excrements recollits i enterrats pels escarabats piloters. Evidentment, hom no ha dut a terme una anàlisi rigorosa de la segregació de nínxols en un grup tan complex, però no hi ha dubte que es poden establir agrupacions funcionalment separades més fàcilment que en el cas dels vertebrats.
Qui es menja a qui
El règim alimentari
La tria de l’aliment pels animals es fa d’una manera instintiva, però eficaç i econòmica. Quan hi ha poc aliment disponible, un animal es veu obligat a menjar qualsevol cosa i, per tant, molts animals mediterranis han reaccionat davant el deteriorament de l’entorn modificant llur règim alimentari. Per exemple, els milans ara són menys caçadors que carronyaires i compensen, d’aquesta manera, l’escassetat creixent de preses. A les illes petites de la Mediterrània, on l’entomofauna és escassa, les sargantanes, naturalment insectívores, esdevenen parcialment herbívores per tal de sobreviure. Però quan hi ha prou aliment, cada espècie desenvolupa una estratègia òptima per a procurar-se menjar; això consisteix a maximitzar l’aportació d’energia per unitat d’esforç destinat a l’alimentació. Aquest tipus d’estratègia, que posa l’eficiència per davant del rendiment energètic brut, correspon a ecosistemes que posseeixen una llarga història i on els animals han pogut acoblar-se bé a les condicions locals.
Una condició bàsica en la tria de l’aliment té molt a veure amb les característiques anatòmiques del sistema digestiu. El fet que els remugants tinguin estómacs especialitzats o els ocells granívors estómacs pedrers trituradors és una causa i una conseqüència alhora d’un règim d’alimentació molt específic. Tot i així, els animals són capaços d’una certa flexibilitat; per exemple, s’estimen més els brots sucosos i tendres de les plantes quan lluquen a la primavera que no pas les tiges bastes i endurides dels pasturatges d’estiu. Els ungulats mediterranis mengen arran de terra, de manera que cerquen els brots acabats de néixer i rebutgen l’herba vella. Naturalment, quan no hi ha res més, també mengen les tiges dures o bé emigren cap a zones millors. Fins i tot els animals mancats d’especialitzacions en l’aparell digestiu presenten unes característiques, especialment les dimensions de la boca, que els fan fer una selecció natural de l’aliment. Per exemple, els animals insectívors, sobretot si no tenen aparell mastegador adequat (és el cas dels ambifis i dels saures) acostumen a triar una grandària òptima de presa, de manera que les xarxes tròfiques no s’estableixen relacionant unes espècies amb unes altres, sinó més aviat relacionant els depredadors amb una determinada categoria de dimensions de les preses. Tot això significa, evidentment, que la major part dels animals han de canviar llurs preferències alimentàries amb l’edat.
La xarxa tròfica
En el bioma del bosc esclerofil·le mediterrani, hi ha tots els nivells tròfics bàsics, excepte en alguns indrets en els quals les comunitat naturals es troben molt degradades. Normalment, els productors són les plantes llenyoses i herbàcies, a les quals cal afegir les algues del sòl, que tenen una importància secundària. Els herbívors són, principalment, insectes, rosegadors i ungulats; els ocells acostumen a practicar una forma especialitzada d’herbivorisme, tot alimentant-se de fruits i llavors. Els aràcnids, molts insectes, els amfibis, petits rèptils i molts ocells insectívors o limícoles integren el nivell dels consumidors primaris. Una bona part dels rèptils, els ocells rapinyaires i els petits mamífers carnívors són, en llur majoria, consumidors secundaris. Els escassos consumidors terciaris són gairebé exclusivament els grans ocells rapinyaires. Els descomponedors consten de grups molt diversos, des de mamífers i ocells que es nodreixen de carronya fins a insectes diminuts i àcars que viuen al sòl, però, en darrer terme, la feina final de descomposició és duta a terme per fongs i bacteris.
L’organització i l’estructura de la xarxa
Heus ací un exemple més detallat de l’organització de la xarxa tròfica en un ambient mediterrani; correspon a una localitat insular de la Mediterrània, en una àrea ocupada per un alzinar molt alterat per la influència humana, que ha substituït els arbres originals per pi blanc (Pinus halepensis) i ha esclarissat el matoll. Els arbres i els arbustos (Erica arborea i diferents espècies de Cistus) constitueixen, juntament amb l’escassa vegetació herbàcia de gramínies, el nivell productor. Aquest nivell envia matèria a altres compartiments de la xarxa: els ocells frugívors i granívors, com ara les tórtores i els trencapinyes, els escassos mamífers herbívors (principalment, ratolins de camp), les tortugues terrestres, els insectes defoliadors i els insectes antòfils dels cists. En darrer terme, les principals sortides d’aquest compartiment van a parar al sòl, on les poden utilitzar els organismes descomponedors, i a l’espècie humana, que les aprofita principalment mitjançant l’extracció de llenya i el pasturatge ocasional.
Els consumidors secundaris són representats, principalment, pels ocells insectívors, els petits rèptils, dues espècies de gecònids, els mamífers insectívors, el més important dels quals és l’eriçó, i els carnívors pròpiament dits, com ara el gat mesquer. En aquest nivell se situen parcialment també alguns ocells majors, com ara els rapinyaires nocturns que es nodreixen de rosegadors herbívors i de rèptils insectívors, o bé els còrvids, que mengen fruits, llavors, insectes i petites tortugues. La sortida d’aquest nivell va al compartiment dels consumidors secundaris i, també, lògicament, al dels descomponedors. Els humans, que formen part de tots els nivells de consum, aprofiten poc aquest últim, principalment a través de la caça. Els consumidors terciaris són solament alguns ocells rapinyaires i algunes serps, com ara la serp blanca (Elaphe scalaris), que s’alimenten sobretot d’ocells petits.
Sobre aquesta estructura bàsica, se superposen diverses xarxes tròfiques menors, acantonades en determinades porcions de la biocenosi major. Per exemple, els troncs dels arbres contenen llur propi nivell de producció, constituït per algues clorofícies i líquens, que reben nutrients no solament de la llum del sol, sinó també de les substàncies dissoltes a l’aigua de la pluja que s’escola pel tronc. Els pugons i altres insectes o àcars viuen dels exsudats de saba i proporcionen aliment a les formigues i a les aranyes corticícoles. Diversos ocells, com els cèrtids, els sítids i els pícids, cerquen dins les esquerdes corticals aquests insectes o les larves dels coleòpters que es refugien a la zona subcortical.
Una altra xarxa tròfica important es troba prop del sòl. Atesa la duresa de l’estiu mediterrani, molts animals cerquen recer i menjar a les esquerdes del sòl o sota les pedres. Allí es reuneix una fauna lapidícola o edàfica, integrada per cucs, nematodes, col·lèmbols i àcars, paneroles, miriàpodes, papaorelles, formigues i cargols. Els animals més grans d’aquesta comunitat són els petits rèptils que, a la conca mediterrània, són principalment gecònids. Malgrat llur règim insectívor, aquests acostumen a cercar aliment a l’exterior, fent servir les pedres soltes o les escorces semidespreses dels arbres únicament com a recer. Ben al contrari, les musaranyes cerquen àvidament sota les pedres els insectes i els cargols que les nodreixen i no romanen gaire temps en un mateix refugi.
Les relacions tròfiques
Ja hem esmentat més amunt que de l’estudi de les xarxes tròfiques semblen sorgir algunes normes generals. L’exigència que les cadenes tròfiques siguin curtes, de tres o quatre baules com a màxim, atesa la ineficiència energètica de la ingestió d’aliment; l’escassetat relativa d’animals que s’alimenten sobre més d’un nivell (omnívors, en el sentit més ampli); la relativa constància del nombre d’espècies que serveixen com a presa, independentment de la complexitat de la xarxa tròfica implicada; l’escassetat relativa de vincles tròfics en comunitats molt complexes, etc., constitueixen generalitzacions, de vegades simples conjectures, que, en cap cas, no han estat comprovades amb tot rigor. Entre aquestes conjectures, algunes es refereixen a l’estructura de l’hàbitat i es podrien estudiar en les comunitats del bioma mediterrani. Per exemple, hom suposa que les característiques de les xarxes tròfiques són influïdes pel nivell de producció de biomassa a l’ecosistema, pel grau de l’estructuració vertical i per la constància o la variabilitat de les condicions ambientals. D’aquesta manera, a les comunitats mediterrànies, amb una producció intermèdia i concentrada a la primavera, amb una estructuració vertical també intermèdia a les àrees de bosc i baixa a les de matoll, i amb una fluctuació regular i intensa de les condicions ambientals, particularment en la quantitat d’aigua disponible, s’haurien de donar cadenes tròfiques curtes, una escassa proporció d’espècies que es nodreixen en diversos nivells i un nombre reduït de connexions per espècie, de manera que cadascuna d’elles tendiria a alimentar-se d’unes quantes, d’una manera força especialitzada.
Aquestes condicions són difícils de comprovar. La literatura científica ha descrit gairebé un centenar i mig de xarxes tròfiques, basades en dades de valor molt desigual, però gairebé sempre insuficient; tanmateix, no hi ha estudis específics que comparin l’estructura general de les xarxes tròfiques mediterrànies amb les d’altres biomes. A la comunitat insular de bosc esclerofil·le que hem descrit més amunt, es poden establir deu cadenes tròfiques diferents, amb una longitud mitjana de 2,8 baules. A la comunitat de matoll mediterrani de Doñana, al sud de la Península Ibèrica, s’han descrit d’una manera sumària més de 250 cadenes tròfiques, amb una longitud mitjana de 3,0 baules. Les dades són força més escasses en el cas de comunitats d’altres regions de clima mediterrani. En una comunitat de mamífers de Xile central, la longitud mitjana de 43 cadenes tròfiques és de 2,1, molt baixa, potser a causa de la limitació taxonòmica.
Un altre tret general de les cadenes tròfiques que hom suposa que canvia amb l’estacionalitat de l’ecosistema és la connectància, o la proporció de relacions existents respecte del total de les possibles. En una xarxa tròfica de N elements, el total de relacions possibles és de (N2-N)/2, però, en realitat, es dóna una fracció molt petita d’aquest nombre, i és més petita com menys estable sigui l’ecosistema. Als ambients de clima mediterrani, per tant, cal esperar xarxes tròfiques de connectància baixa. Per a les xarxes descrites, la connectància oscil·la entre 0,17 i 0,36.
Els valors de longitud mitjana i connectància per a xarxes tròfiques corresponents a ecosistemes estables, com ara els boscos de la zona temperada o els estanys d’aigua dolça, acostumen a ser, efectivament, més alts. Els valors publicats són difícilment comparables, atès que les cadenes tròfiques s’han establert segons diferents criteris, però ens podem referir a comunitats pròximes a les esmentades més amunt i estudiades pels mateixos autors. Per exemple, a l’illa de Menorca, a la Mediterrània occidental, s’obren barrancs arrecerats al sòl de la garriga, on les condicions climàtiques són més estables, la coberta vegetal més densa i l’escassetat d’aigua menor, ja que s’hi canalitza el vessament pluvial. En aquests barrancs, la llargària mitjana de les cadenes tròfiques és clarament major, de 3,9 baules, i en comunitats d’aigua dolça de la mateixa illa, encara més estables, aquesta llargària assoleix 5,1 baules. Els valors de la connectància també són més alts, entre 0,19 i 0,49.
Aquests resultats no són pas concloents, atès que les característiques generals de les xarxes tròfiques canvien en el mateix sentit quan augmenta la complexitat, i les comunitats estables acostumen a ésser més complexes. No obstant això, permeten de cercar una explicació de les possibles diferències. És evident que les cadenes curtes augmenten l’eficiència en la circulació de l’energia, ja que, en cada baula de la cadena se’n perd la major part. Les conclusions apuntades concorden amb el fet que, en un clima mediterrani, on els recursos escassegen cada estiu, sigui útil d’alimentar-se amb eficiència i constituir cadenes tròfiques curtes. De fet, els animals que viuen en aquest ambient es nodreixen amb molta eficiència gràcies al canvi periòdic de dieta: sempre es procuren aliments que es trobin en el moment àlgid de producció.
Una característica força general de les cadenes tròfiques al bioma mediterrani és el paper clau que hi tenen els rèptils. Com que alguns, especialment les serps, són capaços de nodrir-se de mamífers o d’ocells i, al seu torn, acostumen a ser caçats per ocells i mamífers com a presa, no és estrany que formin part de cadenes tròfiques particularment llargues. En aquestes cadenes, el vèrtex sol estar ocupat per un depredador estenòfag, generalment un ocell rapinyaire, amb un predomini de rèptils a la seva dieta. És el cas de l’àguila marcenca (Circaetus gallicus) a la Mediterrània, el serpentari (Sagittarius serpentarius) a Sud-àfrica, de l’Aquila audax a Austràlia, l’aligot Buteo jamaicensis a Califòrnia o Geranoetus melanoleucus a Xile, malgrat que aquestes últimes espècies no s’especialitzen en la caça de rèptils. La baula inicial és, evidentment, la vegetació, però de vegades se superposen les cadenes tròfiques epigees amb les dels descomponedors, formades per la fauna del sòl. D’aquesta manera poden donar-se cadenes excepcionalment llargues, que s’inicien en els bacteris i continuen amb els nematodes, els àcars, els col·lèmbols, els pseudo-escorpins, els caràbids, els ocells insectívors, els llangardaixos, les serps i els ocells rapinyaires.
Migradors i sedentaris
Els organismes que viuen a les garrigues mediterrànies són sotmesos a la privació d’aigua durant l’època estiuenca. Si aquesta mancança fos permanent, caldria que adoptessin mecanismes extrems per a pal·liar-la, tal com passa en el cas de les espècies desèrtiques, o bé serien eliminades per manca d’adaptació. La periodicitat de la sequera permet que l’activitat d’animals i plantes adquireixi un ritme també periòdic, i que la principal activitat nutritiva i reproductòria coincideixi amb el desvetllament del repòs hivernal, en què la disponibilitat d’aigua és l’adequada.
Molts animals adapten llur ritme al de la natura mediterrània; d’altres han optat pel camí alternatiu de traslladar-se d’acord amb l’ocasió i recorren a la migració per tal de trobar sempre l’ambient més adequat. Aquest tret permet de dividir els animals del bioma mediterrani en migradors i sedentaris. No obstant això, la diferència només és de grau: tots els animals es traslladen a la recerca de protecció o d’aliment, i molts ho fan d’una manera periòdica; però hom parla d’animals migratoris quan llurs recorreguts es repeteixen si fa no fa al cap d’un cicle, i quan abracen distàncies geogràficament significatives que els comporta un desplaçament important.
Les grans migracions dels ocells
Els animals que fan grans migracions són, lògicament, els ocells. Llur capacitat de vol els permet de fer desplaçaments molt dilatats que, en casos extrems, van des de l’Àrtida fins a l’Antàrtida. Sense arribar a fer els considerables recorreguts dels ocells oceànics, molts ocells aprofiten l’oposició de les estacions en els dos hemisferis terrestres per a poder estar sempre en indrets rics en aliment; així, durant l’estiu septentrional, es desplacen cap al nord per a reproduir-se, i durant l’hivern canvien d’hemisferi i poden aprofitar l’estiu meridional. Les migracions no afecten només els països temperats o bé els freds, sinó que també es donen en ocells tropicals i subtropicals, lluny de les àrees de clima mediterrani.
L’estratègia migratòria
Molts ocells fan migracions àmplies, però menors, que no requereixen canviar d’hemisferi. Aquests ocells simplement pretenen fugir dels rigors de l’hivern àrtic i es refugien en zones més meridionals amb hiverns més suaus. La conca mediterrània és una d’aquestes àrees de característiques tèrmiques tolerables, que serveix de zona d’hivernada per a força espècies procedents del nord. Durant l’hivern, es barregen, en aquesta regió, els ocells que hi vénen a hivernar des del nord d’Europa, les espècies sedentàries, que hi romanen, i les migrants parcials, que normalment cerquen indrets més càlids a l’Àfrica, però que solament part de la població hi migra, mentre que la majoria romanen a la conca mediterrània. A aquests grups cal afegir, en èpoques de migració, els ocells de pas, que reposen provisionalment a l’àmbit mediterrani abans de continuar llur viatge cap al sud a la tardor i cap al nord a la primavera. A la primavera també s’hi dóna una barreja d’espècies, amb diferents conductes migratòries, en aquest cas a causa del retorn als indrets de cria al nord.
Fora de la conca mediterrània, les grans migracions són menys importants. En el cas d’Austràlia i de Sud-àfrica són poc importants, ja que no hi ha terres més al sud on hom pugui anar durant la primavera (tret de l’Antàrtida, que no serveix per a aquest propòsit). Austràlia podria servir de punt de destinació per als ocells nord-asiàtics terrestres (de fet, molts ocells oceànics hi hivernen), però acostumen a aturar-se, si bé no tots, a Indo-xina. Evidentment, Sud-àfrica és el lloc de destinació d’una part de l’avifauna eurasiàtica durant l’hivern septentrional (per exemple, moltes cigonyes de l’Europa oriental hi hivernen), però només una dotzena d’espècies, i algunes més que no niuen a la conca mediterrània, assoleixen la regió del Cap; en aquest sentit, la regió forma part del sistema de migració paleàrtic.
La regió mediterrània xilena tampoc no es presta a les grans migracions. Ultra el problema indicat de manca de terres a latituds meridionals, els ocells d’aquesta zona tenen la serralada dels Andes ben a prop, que els permet de substituir el canvi de latitud per un canvi d’altitud, molt més breu i menys arriscat; els ocells marins procedents dels arxipèlags del sud fan servir, evidentment, la regió mediterrània xilena de zona de pas, i per a la gavina andina és una de les zones d’arribada durant la primavera austral. Ben al contrari, la regió californiana es troba dins una de les sis grans rutes de migració del sistema neàrtic, malgrat que és menys utilitzada com a àrea de destinació que no pas com a zona de pas. Força espècies procedents del Pacífic nord, d’Alaska i fins i tot de l’altra banda de l’estret de Bering hi recalen, no a la garriga sinó a les llacunes i als aiguamolls veïns, però la majoria continua el seu viatge fins a Mèxic i l’Amèrica Central.
En canvi, la conca mediterrània ofereix característiques molt adequades per a mantenir una elevada varietat estacional d’ocells. A la suavitat tèrmica dels seus hiverns s’afegeix el fet que a la part nord se situen grans penínsules que es projecten cap al sud i que s’acosten al litoral africà. Aquestes penínsules constitueixen punts obligats de pas per als ocells migradors, que després de sobrevolar-les només cal que travessin braços de mar força estrets. La facilitat de pas es fa més evident amb l’existència de nombroses illes a la Mediterrània, que fan de punts de repòs de la travessa. D’altra banda, hi ha vuit àrees pantanoses principals i centenars d’àrees pantanoses secundàries, amb una elevada producció d’aliment a diversos països de la zona, des de les maresmes del Guadalquivir fins al delta del Nil. Aquestes àrees permeten als ocells migratoris, especialment els limícoles, els anàtids i els ardèids, d’alimentar-se i descansar durant el viatge, i tot sovint esdevenen indrets de destinació definitius per a la hivernada. Per totes aquestes raons, prop de la meitat de l’avifauna europea migra, totalment o parcialment, a través de la Mediterrània.
Les rutes migratòries
Així doncs, les grans rutes d’aquesta mar segueixen el camí de les principals penínsules. Un contingent important circula per l’estret de Gibraltar, un altre encara més gran per la mar Negra, la costa d’Anatòlia, la costa de Síria, el Líban, Israel, el Sinaí i penetra a l’Àfrica pel delta del Nil; el tercer, menys important, segueix la Península Itàlica o travessa el mar amb l’ajut de les illes de la zona: Sicília, Còrsega, Sardenya i les illes menors. El primer i el tercer corredor són útils per a l’avifauna europea i acaben a l’Àfrica occidental, sense passar a l’hemisferi sud, malgrat que alguns dels ocells que els utilitzen hivernen a totes dues bandes de l’equador. El segon, i principal, afecta no sols els ocells migratoris d’Europa, sinó també els de gran part d’Àsia i acaba a l’extrem sud de l’Àfrica. Les rutes esmentades són força difuses i es combinen amb camins més curts, recorreguts per espècies de migració breu o per migradors parcials, que en fan amplis corredors amb serrells laterals.
La mortaldat dels ocells durant la migració és molt elevada: potser un terç dels individus que comencen la migració moren entre el viatge d’anada i el de tornada, i aquest nombre és de molts milions l’any. L’agent principal d’aquestes morts és el temps meteorològic. Una tempesta o un huracà poden fer morir milions d’ocells en vol; potser aquesta és la raó per la qual la ruta de l’Àsia oriental s’atura tot sovint a Indo-xina: el mar de la Xina meridional és un centre productor de ciclons que fa difícil d’arribar fins a Austràlia. De fet, el flux d’ocells migratoris a l’Àsia és, principalment, de l’est a l’oest, i això potser es deu a l’existència de barreres, com ara les serralades de l’Àsia central o els centres ciclònics del Pacífic occidental. A les causes meteorològiques cal afegir la predació que exerceixen nombroses espècies, entre elles la humana, que sotgen el pas de la migració.
Cal que ens preguntem per què els ocells han desenvolupat un sistema tan costós en termes de despesa energètica i de supervivència per a aconseguir més aliment. És evident que la conducta migratòria és una adquisició recent. És probable que, en la forma actual, no existeixi més que en l’Holocè, atès que la presència del casquet glacial degué alterar o anul·lar els possibles comportaments anteriors d’aquesta mena. Dotze mil anys poden semblar un termini força breu per a la implantació d’una conducta molt complexa i generalitzada, però no hi ha cap dubte que n’hi ha hagut prou. A més, ha permès a d’altres ocells de modificar la seva pauta de migració per tal d’aprofitar la de la resta d’espècies, tal com ha passat en alguns ocells rapinyaires que s’alimenten d’espècies migradores.
L’arribada dels ocells de pas
A tota la conca mediterrània, l’arribada dels ocells en el seu viatge d’anada o de tornada constitueix un dels moments culminants del ritme anual. Els ocells que arriben a la primavera són rebuts com a presagi de bon temps, i els que se’n van a la tardor tot sovint són esperats com a peces de caça. A mitjan gener, de vegades abans, arriben les primeres orenetes (Hirundo rustica) a la riba de la Mediterrània i llur tornada és eglaonada fins ben entrat el mes d’abril, en què ja arriben a l’Europa central. Aquesta espècie és tan coneguda i esperada que sovint es fa servir com a element d’observació fenològica pels meteoròlegs. Hi van darrere els ocells insectívors, que esperen que es desvetlli l’entomofauna mediterrània, i tanca el cicle la cigonya, que s’instal·la quan ja pot trobar aliment a les tolles o als camps, o bé prossegueix el seu camí cap al nord.
A la tardor, els ocells insectívors constitueixen els vols més nombrosos: tords, pit-roigs, titelles, etc., molts d’ells emigrants parcials. Els humans sotgen l’arribada dels ànecs, normalment inassolibles ja que volen molt alt, però que es reuneixen a les zones de maresma i proporcionen als habitants dels encontorns una caça abundant. Els estornells envaeixen el cel del sud d’Europa ben entrada la tardor, formen bandades molt grans que cobreixen completament els indrets on es posen. Normalment, els ocells tendeixen a evitar els grans sistemes muntanyosos i, per exemple, envolten els Alps per l’est i l’oest per no haver de sobrevolar-los; en qualsevol cas, quan els travessen, s’estimen més de fer servir els ports i les collades de menys altitud. La canalització de la migració pels ports de muntanya ha estat aprofitada des de temps molt antic pels humans de la Mediterrània, que sotgen, a la muntanya, els dies de la migració. La caça del tudó (Columba palumbus), per exemple, ha donat lloc a una forma de caça molt organitzada, amb espieres a les muntanyes que fan servir any rere any els mateixos caçadors i que es passen de pares a fills.
L’espècie humana no és l’única caçadora que espera els ocells migratoris; com s’ha esmentat més amunt, altres depredadors comparteixen aquesta espera. Un dels més importants és, precisament, un endemisme de la conca mediterrània, el falcó de la reina (Falco eleonorae). Aquest rapinyaire roman a la zona durant la tardor i és una de les poques espècies d’ocells que cria en aquesta estació; d’aquesta manera, pot nodrir els seus polls amb les abundants preses que li proporciona la migració de les espècies nòrdiques. Després, al final de la tardor, es dirigeix cap al sud de l’Àfrica per hivernar-hi. Aquest falcó vigila el corredor occidental de migració, mentre que un congènere seu, el falcó cendrós (Falco concolor) controla la gran ruta de la Mediterrània oriental.
Altres desplaçaments migratoris
El terme migració sempre s’associa als ocells i als moviments de gran abast i de periodicitat estacional. Lògicament, hom no pensa en els desplaçaments ocasionals, que poden ser importants, però que no obeeixen a cap ritme, ni en els moviments periòdics d’escassa amplitud, ni tampoc en els llargs recorreguts periòdics que duen a terme altres grups diferents d’animals. Tanmateix, aquests moviments tenen una importància i una significació ecològica semblants a la de les migracions enteses en el sentit habitual. La conseqüència principal n’és augmentar la disponibilitat de recursos tròfics i, per tant, contribuir a l’estabilitat de les poblacions. En efecte, la teoria ecològica prediu que les poblacions que poden canviar llur localització espacial aconsegueixen mantenir una certa seguretat davant qualsevol tipus de catàstrofe demogràfica. Atesa l’heterogeneïtat ambiental derivada de l’estructura en mosaic dels paisatges mediterranis, aquestes zones són òptimes perquè una espècie hi pugui diversificar les residències a través de desplaçaments periòdics o irregulars, que poden abastar milers de quilòmetres, com el cas dels ocells migratoris, o bé es poden reduir a uns quants mil·límetres, com en el cas de la microfauna del sòl.
Molts dels animals no sedentaris de les àrees mediterrànies fan moviments estacionals de recorregut curt. Un component important de l’avifauna del “mallee” australià, integrat majoritàriament per psitàcids de dimensions reduïdes, duu a terme desplaçaments graduals entre el nord i el sud del continent i segueix els canvis estacionals de la vegetació, especialment la producció de llavors. A la resta d’àrees amb clima mediterrani, la proximitat de sistemes muntanyosos més o menys elevats permet d’aprofitar la variabilitat de condicions climàtiques inherent a l’altitud. Alguns animals porten a terme moviments estacionals de tipus transhumant entre les terres altes i les baixes, o entre àrees veïnes que tenen èpoques de producció màxima desfasades. Entre els ocells transhumants de la regió mediterrània xilena, hi ha falcillots, papamosques, orenetes i diversos pinsans, en general ocells insectívors parcialment migratoris, que cerquen l’aliment a la muntanya o a la plana (també fan migracions horitzontals), d’acord amb l’època d’activació de les preses.
Les migracions d’insectes alats
Tal com hem esmentat més amunt, les migracions pròpiament dites semblen privatives dels ocells. De fet, però, hi ha animals que pertanyen a altres grups que duen a terme moviments periòdics de gran amplitud. Entre els animals terrestres de les zones de clima mediterrani, el més conegut n’és un insecte, la papallona monarca (Danaus plexippus). Aquesta espècie americana ha estat profundament estudiada pels moviments estacionals que efectua entre Canadà i Mèxic, on pot recórrer milers de quilòmetres. Encara que les rutes principals de migració travessen l’interior dels Estats Units i Mèxic, una bona part de les papallones procedents del vessant pacífic de l’Amèrica del Nord hiverna a les costes càlides de l’Alta i la Baixa Califòrnia, on prefereixen les zones arrecerades i amb arbres més que no pas les zones de garriga. Molt sovint, nombrosos exemplars d’aquesta espècie, que també cria a les Illes Canàries, són desviats pels vents i apareixen a Europa, i sobre la base de les illes del Pacífic han anat ampliant llur àrea de distribució fins a arribar a Austràlia. D’altra banda, aquest animal té la seva contrapartida europea en un congènere també mediterrani, la papallona tigre (Danaus chrysippus), pròpia de la Mediterrània oriental, i que també emigra d’Europa a l’Àfrica, si bé els seus moviments no han estat gaire estudiats; altres espècies de papallones duen a terme migracions menors entre la Mediterrània i l’Europa central.
Les costes de la mar Mediterrània, que representen les formes típiques d’aquest bioma, són l’escenari de moviments estacionals de tota mena. Les serralades muntanyoses estan disposades amb orientacions molt diverses, en general paral·leles a les costes veïnes, de manera que els moviments verticals tenen moltes possibilitats. Exemples de desplaçaments periòdics d’aquest tipus i d’altres es donen en molts grups d’animals que habiten a la conca. Entre els insectes, no és pas la papallona tigre l’única espècie migrant; altres papallones (per exemple, la Vanessa cardui) realitzen viatges a les zones altes a la primeria de l’estiu, com també altres insectes, com la marieta comuna (Coccinella septempunctata).
Al sud de Califòrnia, les condicions són molt semblants a les de la conca mediterrània europea. Les muntanyes pròximes i els arxipèlags costaners han afavorit l’aparició d’endemismes. Aquests, lògicament, presenten una mobilitat baixa, però les espècies de distribució més àmplia inclouen grans viatgers. Ja hem esmentat més amunt el cas espectacular de la papallona monarca, que el mateix individu viatja més de 6-000 km; altres insectes amb desplaçaments estacionals importants són les marietes, que tenen un comportament similar en molts sistemes muntanyosos de l’hemisferi nord: es congreguen als cims a l’inici de l’estiu i en baixen a la tardor. Una espècie californiana, Hippodamia convergens, es reuneix en grans grups de milions d’individus abans i després del seu viatge anyal.
Les migracions de peixos i mamífers
Els animals no voladors tenen menys possibilitats de fer grans desplaçaments. No obstant això, els grans herbívors segueixen la fenologia de la vegetació i es traslladen a les zones en què hi ha brots nous. Aquests moviments es palesen sobretot a les àrees muntanyoses, on consisteixen en canvis d’altitud. Diferents espècies d’ovins salvatges s’han diferenciat a les muntanyes mediterrànies (cabra salvatge, mufló, pasang, etc.); pugen a les pastures d’estiu quan la neu es retira i tornen a les valls amb les primeres nevades. Aquestes migracions verticals també es donen en animals no muntanyesos, més aviat específics del bosc mediterrani, com ara el senglar o la daina, i, a la regió californiana, el cérvol mul (Odocoileus hemionus), una espècie forestal que sovinteja a les àrees de garriga. Probablement, la transhumància del bestiar domèstic s’originà a la prehistòria a partir de l’observació dels moviments de les espècies salvatges.
A Sud-àfrica, hom observava migracions territorials d’aquesta mena en nombrosos bòvids. Un antílop molt simbòlic per a aquest país, la gasela saltadora (Antidorcas marsupialis) constituïa antigament ramats de milions d’individus, que feien desplaçaments estacionals massius a la recerca de pastures idònies pel Karoo i les regions properes; tanmateix, la caça legal i il·legal n’ha reduït dràsticament els efectius, que ara es troben protegits en alguns parcs i reserves. Es tracta d’una història semblant a la del conegut bisó americà.
Altres animals alats fan llargues migracions; per exemple, diferents espècies de rat-penats es retiren durant l’hivern en coves arrecerades que els serveixen de refugi, i tornen a dispersar-se a la primavera, quan l’entomofauna que els nodreix ja és prou important. Les migracions dels rat-penats han estat estudiades mitjançant anellaments, com en el cas dels ocells, i hom ha deduït que aquestes migracions són importants, sovint amb recorreguts de molts centenars de quilòmetres. Una conducta similar presenten algunes espècies de rat-penats de la regió mediterrània xilena, mentre que d’altres (sobretot Lasiurus borealis i Lasiurus cinereus) realitzen veritables migracions cap al nord. Els mamífers no voladors tenen una menor capacitat de desplaçament, i la majoria no depenen tant dels insectes o de les llavors. Els grans herbívors són espècies sovint forànies, introduïdes des de la regió mediterrània europea, i presenten hàbits transhumants. Així, el cérvol comú, la daina, la cabra salvatge o el senglar, sobretot aquest darrer, efectuen moviments estacionals cap a o des de la muntanya.
Entre els vertebrats desproveïts d’ales, potser les migracions més famoses són les de les anguiles (Anguilla), uns animals que, si bé durant la fase juvenil no pertanyen al bioma mediterrani, sí que s’hi integren a l’edat adulta, quan remunten els rius de les contrades riberenques. Cal destacar que els humans han interferit decisivament en aquesta mena de migració, ja que han tallat, per mitjà de preses i embassaments, la possibilitat d’assolir el curs alt dels rius. Aquest fenomen, que es dóna a tota la Mediterrània, ha comportat la pràctica desaparició de l’anguila a l’interior de diversos països, entre els quals cal esmentar Espanya.
Els animals sedentaris
Són qualificats com a sedentaris els animals que es desplacen en petites distàncies, per oposició a les espècies més mòbils, si bé aquesta definició és força imprecisa. En general, els animals sedentaris acostumen a romandre en llur territori, quan existeix, o a fer desplaçaments erràtics i irregulars. Molts d’ells es traslladen amb una certa periodicitat, fins i tot d’una manera vinculada al cicle estacional. Força coleòpters voladors realitzen cada dia viatges sincronitzats entre les zones d’alimentació i de reproducció; els cargols petits, tan abundants a la garriga mediterrània, tenen tendència a pujar, durant l’època estiuenca, per les tiges de les plantes i es concentren a les branques superiors i a les inflorescències seques, molt sovint entapissades per centenars d’individus. Les mosques i els mosquits que cerquen les zones més humides s’acumulen per milers a les parets fresques i ombrívoles d’algun barranc i esdevenen actius amb l’arribada del crepuscle, en què la calor minva. La fauna del sòl presenta curts moviments d’ascens i descens que segueixen les oscil·lacions tèrmiques quotidianes. En general, aquests moviments d’escàs recorregut estan lligats als ritmes nicthemerals. Els moviments de caire estacional es vinculen, en canvi, a les fases anyals d’activitat i repòs, que depenen de l’alternança entre període humit i període eixut característica de l’àmbit mediterrani. Els animals van a la recerca de refugis adequats per a hivernar-hi o per a passar-hi l’estiu, segons els casos.
El ritme de la vida animal
Els animals se solen adaptar als canvis a la natura tot prevenint-los, si és possible; és a dir, si aquests canvis es presenten amb una certa regularitat. Pel que fa a la fauna mediterrània, com per a la de les altres zones, els canvis regulars més importants són els derivats de l’alternança de dies i nits, que donen lloc a la manifestació de ritmes circadiaris. També són força importants els ritmes derivats de la successió de les estacions, i són molt més complexos.
A una escala temporal intermèdia entre els cicles curts anuals i les grans oscil·lacions climàtiques del Quaternari, se succeeixen al bioma mediterrani alguns canvis irregulars, que es repeteixen amb seguretat, però sense periodicitat. Aquests canvis generalment són catastròfics, i a diferència de les variacions climàtiques tenen un caràcter local. Durant molt de temps, un indret determinat en pot romandre al marge, però tard o d’hora, després d’un període que no ultrapassa mai uns quants segles, hi arriben. Atès el caràcter variable dels períodes de recurrència, la natura no ha pogut preveure aquestes catàstrofes mitjançant l’adquisició de ritmes endògens que permetin a la flora i a la fauna d’avançar-s’hi, però la seguretat que en un moment o un altre es presentaran, ha provocat una certa pressió selectiva, a conseqüència de la qual predominen en l’ambient mediterrani formes pre-adaptades a aquests esdeveniments catastròfics.
Els ritmes quotidians
El recurs de la vida nocturna representa, en els animals terrestres, una oportunitat de protegir-se davant de dos tipus de perills: la depredació per part d’animals el món sensorial dels quals es regeix per la vista, com ara els ocells, i la dessecació produïda per la calor solar. D’altra banda, l’exclusió competitiva fa que molts animals mantinguin una activitat nocturna i que ocupin nínxols inaccessibles als animals diürns.
A les zones de clima mediterrani es donen tots aquests factors. Els animals diürns són principalment ocells, rèptils i insectes florícoles, mentre que els nocturns són els que posseeixen un entorn sensorial olfactiu (mamífers) o que depenen de la humitat en un grau major (insectes epigeus i amfibis); els mol·luscs no tenen preferències i entren en activitat quan hi ha prou humitat. Però la necessitat d’aigua durant la sequera estiuenca indueix molts animals a canviar les pautes d’activitat, i per tant el caràcter de diürns o nocturns depèn de les èpoques de l’any.
A més, el màxim diürn d’activitat es desplaça dins el dia o la nit, segons quina en sigui la durada i segons les condicions tèrmiques i d’humitat. A les latituds més septentrionals, els rellotges interns dels animals són relativament fixos, fins al punt que els humans els han fet servir des de la prehistòria com a complement del rellotge celest per a mesurar el recorregut del dia. A les regions mediterrànies, els ritmes biològics són menys estables, però tot i així presenten unes pautes definides.
Quan surt el sol, són els ocells, generalment els petits passeriformes, els qui comencen l’activitat; les espècies que dormen agrupades als arbres comencen el dia amb crides d’integració social i envaeixen les capçades amb els seus refilets. A Amèrica, les necessitats metabòliques obliguen els colibrís a iniciar la recerca de nèctar així que el dia clareja. En general, els ocells de dimensions petites tenen una taxa metabòlica més elevada i necessiten alimentar-se més d’hora. La resta dels ocells diürns comença l’activitat després que el sol ja ha sortit.
Pel que fa als rèptils, la sortida dels refugis té lloc quan el sol comença a escalfar, i els dies núvols i freds acostumen a romandre inactius. Els mesos més càlids de l’estiu, tanmateix, poden tornar a llurs refugis després d’una ràpida caça d’insectes, abans que la calor esdevingui insuportable i, en aquest cas, poden tornar a sortir durant el vespre o la nit. Poc abans que es comencin a moure, han començat a fer-ho molts insectes epigeus, els quals precedeixen les espècies florícoles i voladores, que surten quan ja és el pic del dia.
Els mamífers petits presenten una activitat discontínua en trams curts, que s’alternen amb el repòs al refugi, i que es manté durant la nit. Naturalment, si la calor o el fred són massa forts, romandran amagats. Les musaranyes no es poden quedar gaire temps en repòs i gairebé constantment cerquen menjar, amb predomini de l’activitat nocturna. Els grans herbívors s’estimen més de menjar de bon matí, en què el sol ha evaporat la rosada, i cap a mig matí es retiren a l’ombra de matolls o roques, mentre remuguen la ingesta. Abans del vespre tornen a menjar, i durant la nit acostumen a romandre en repòs.
Durant la tarda, i especialment al vespre, quan el sol comença a pondre’s, els insectes es retiren, generalment cercant refugi sota les pedres, mentre que les espècies voladores romanen quietes sobre les plantes. Altres grups, principalment dípters i petits lepidòpters que no es nodreixen de nèctar i que requereixen un ambient més humit, tenen una activitat predominantment vespertina o nocturna, i surten quan el sol es pon. Llavors entren també en activitat els ocells especialitzats en aquest tipus de dieta, com ara els caprimulgiformes, com també els rat-penats insectívors, el grup crepuscular més conegut. La caça de mosquits i arnes també la fan espècies que han esdevingut comensals de l’espècie humana, com ara els gecònids mediterranis, i, a causa d’això, la seva pauta d’activitat mostra un màxim a primera hora de la nit, en què els insectes voladors es reuneixen sota els llums dels carrers i els edificis.
A la nit, molts mamífers presenten un màxim d’activitat. Per exemple, les musaranyes i els ratolins, tot i que també es mantenen actius durant el dia, o alguns rat-penats, que continuen llur activitat crepuscular. Els registres de l’activitat de diversos rosegadors de la regió del Cap revelen patrons molt variats; la majoria són nocturns, fins i tot quan viuen habitualment sota terra. Hom creia que els petits carnívors (mustèlids, vivèrrids, etc.) caçaven principalment de nit, però observacions recents amb radiotelemetria indiquen que una bona part de la seva activitat és diürna.
Els estrigiformes són uns ocells especialitzats en la caça nocturna i pràcticament no volen de dia, però no són pas els únics; altres espècies mediterrànies, com ara el torlit (Burhimus oedicnemus), es mouen sobretot a la nit. Els rèptils nocturns són molt escassos, a causa de les seves necessitats tèrmiques, mentre que als amfibis terrestres (bufònids, discoglòssids, leptodactílids, etc.), les necessitats d’humitat els imposen caçar exclusivament a la nit. L’activitat reproductora en aquests animals també té lloc de nit. Entre els invertebrats, són nocturns els insectes lapidícoles, com ara els tenebriònids i els caràbids, els cucs i la majoria de les aranyes i dels cargols.
Per últim, abans de sortir el sol, hom arriba a un mínim general en l’activitat animal; malgrat que alguns grups romanen actius, la fauna sembla que reposi abans d’iniciar el nou dia. A aquesta hora comencen les patrulles de caça els grans carnívors, el linx, el llop o el puma. Una mica més tard, quan el sol ja comença a sortir, el cicle recomença.
Els ritmes estacionals
Ja hem comentat més amunt, en parlar dels moviments migratoris, que la fauna mediterrània experimenta canvis periòdics anuals. En realitat, al bioma mediterrani, els dos recursos climàtics bàsics, temperatura i humitat, canvien tot fent oscil·lacions al llarg de l’any, però d’una manera oposada: mentre que el màxim tèrmic té lloc a l’estiu i el mínim a l’hivern, amb la humitat passa tot el contrari, de manera que en la proximitat dels solsticis les condicions no són gaire bones per a la fauna. Per tant, les millors èpoques de l’any són les corresponents als equinoccis, en què la temperatura és força alta, però la sequera no és excessiva. Aquesta circumstància modifica la corba d’activitat anual de moltes espècies, que és bimodal, amb dos màxims, en lloc de ser unimodal, com la que presenten els animals dels boscos temperats i freds.
La pauta indicada no és aplicable a cada espècie, sinó més aviat al conjunt de la fauna. Per a les espècies en particular, és evident que l’època de màxima activitat anual dependrà dels recursos que necessitin i del grau d’adaptació a la seva aparició, si bé moltes espècies posseeixen ritmes endògens que es manifesten independentment de les condicions ambientals. Els cicles anuals en els animals mediterranis es diferencien, principalment, per la intensitat i la situació dels períodes de repòs: la hivernació gairebé mai no és completa i es pot reduir a unes quantes setmanes de repòs en un refugi, mentre que la diapausa estival, que no es presenta en espècies de latituds més elevades, és freqüent a l’àmbit mediterrani i acostuma a ser més intensa i prolongada que no pas el repòs hivernal. Però l’activitat global dels animals no és l’únic element que en defineix el cicle anual: la gametogènesi, les diferents fases de la reproducció, l’alimentació i l’acumulació de reserves i, en el seu cas, la fase de diapausa, la muda o la migració, en són altres trets, cadascun dels quals amb la seva pròpia pauta temporal, cosa que ofereix a les espècies mediterrànies nombroses oportunitats de segregació i d’aprofitament dels recursos.
Naturalment, també hi ha diferències en el patró anyal d’activitat entre les diferents regions. A la conca mediterrània i a Califòrnia, l’activitat és màxima a la primavera, mentre que a Xile central s’estén fins a la primeria de l’estiu i a Austràlia fins ben entrada aquesta estació. De fet, la flora del “mallee” australià, d’origen tropical, sembla conservar les pautes de desenvolupament anual dels seus antecessors terciaris, i moltes espècies hi broten en ple estiu, cosa que obliga els animals a alimentar-se quan la calor és forta. L’activitat de la fauna del sud d’Austràlia és, doncs, regulada en gran part per les disponibilitats d’humitat. D’altra banda, a les dues zones de l’hemisferi nord hi ha màxims hivernals secundaris de pluja que són molt més reduïts que no pas a l’hemisferi sud.
L’activitat primaveral
El cicle comença cada any quan es desvetllen de llur letargia hivernal els animals que reposen durant aquesta estació. Les plantes, moltes de les quals produeixen brots i flors durant l’hivern, són les primeres a reactivar-se, un cop desvetllat el seu ritme endogen amb l’augment de les hores diàries de llum. Pel que fa als animals, en canvi, són generalment les condicions externes, principalment la temperatura, les causants de la reactivació; no obstant això, molts animals també regulen llur activitat, especialment el període reproductor, per mitjà del fotoperíode, de manera que, tot canviant-lo, es pot avançar o retardar l’activitat sexual. Mitjançant aquest mecanisme de sincronització, els mamífers, per exemple, poden iniciar la gestació de manera que el part s’esdevingui a la primavera, en què hi ha abundor de recursos per a nodrir les cries, o els pugons, els himenòpters, etc., que produeixen els ous de diapausa abans de l’hivern per tal que les femelles partenogenètiques apareguin a la primavera i puguin reiniciar el cicle. Els cicles més marcats i estables són els de les papallones, ja que les seves erugues depenen de plantes específiques, i cal que la reproducció i la posta dels adults s’adapti al seu rebrot. Segons les espècies i les localitats, el nombre de generacions anyals canvia i es manté sempre relacionat amb l’existència d’aliment.
A més de les espècies hivernals, les primeres flors apareixen al gener (al juliol a l’hemisferi sud), i al mes següent hom comença a veure els insectes, sobretot les espècies nocturnes, que necessiten més humitat. Així doncs, es mou la fauna epigea de miriàpodes i tenebriònids i, abans que no arribi la primavera, és acompanyada per molts altres coleòpters, que ja es poden menjar les fulles noves de les plantes. Els coleòpters florícoles, els himenòpters nectarívors i les papallones apareixen, lògicament, quan les flors són més abundants, és a dir al març i a la primeria d’abril (setembre i octubre per a l’hemisferi sud). En aquell moment, els ocells migradors ja han arribat i els insectívors inicien llur activitat reproductora, alimentant-se intensament i preparant llur posta.
Al març també entren en moviment els petits mamífers, la majoria dels quals han conservat alguna activitat durant l’hivern. Els insectívors i els rosegadors disposen de prou aliment per a començar a criar. El màxim secundari de pluges primaverals permet, a més, de continuar l’activitat hivernal dels cargols, i d’iniciar la dels amfibis que, en general, han acabat la posta un o dos mesos abans de l’estiu. Els grans mamífers herbívors poden aprofitar l’abundor primaveral de biomassa per a nodrir-se i alletar les cries, que han passat tot l’hivern gestant-se.
Durant la primavera avançada, apareixen els animals que ocupen nivells superiors en la cadena tròfica: serps, rapinyaires, copròfags, etc.; tots ells assoleixen el màxim durant l’estiu. L’existència de ventregades recents de mamífers i de covades d’ocells a la darreria de la primavera augmenta, també, la presència i l’activitat dels petits mamífers carnívors.
L’activitat estival
Al començament de l’estiu, l’època de floració intensa ja ha passat, si bé moltes espècies floreixen durant aquesta estació o durant les següents. Amb això disminueix molt l’abundor de coleòpters florícoles i de nectarívors, però moltes papallones romanen actives durant l’estiu, després del màxim maig-juny (novembre-desembre a l’hemisferi sud), i entre els himenòpters augmenta l’activitat dels granívors, principalment formigues, i dels que duen a terme postes paràsites. Amb l’abundor de fems procedents dels grans herbívors, augmenta notablement l’activitat dels escarabats copròfags (piloters), que recullen reserves d’aliment. Altres escarabats defoliadors (Melolontha, Polyphylla, etc.) presenten, també, un màxim d’activitat al voltant del solstici d’estiu. També és estival el màxim d’activitat dels petits ortòpters, capaços de nodrir-se de fulles relativament endurides, o de molts hemípters (cicàcids, pentastòmids, etc.), que s’alimenten xuclant saba. Els artròpodes carnívors, com els caràbids (que han estat precedits per espècies fitòfagues de la mateixa família), els cicindèlids o les aranyes de terra (licòsids), també són abundants a l’estiu, en què l’entomocenosi mediterrània assoleix encara prou varietat i abundor. Ben al contrari, els cargols, enfrontats al començament de la sequera anual, segellen la seva closca i es refugien sota les pedres o al capdamunt de les tiges especialment herbàcies.
Pel que fa als vertebrats, els amfibis es troben limitats també per la manca d’aigua; durant l’estiu, surten a caçar sols de nit, i si fa calor, també a la nit es queden als refugis. Com per als gastròpodes, l’estiu és per als amfibis l’època de diapausa, i algunes espècies romanen soterrades durant unes quantes setmanes. A Austràlia, dues espècies de leptodactílids (Limnodynastes dumerili i Neobatrachus pictus) passen l’estiu en cavitats subterrànies segellades amb fang per tal de mantenir la humitat. A la conca mediterrània, altres espècies adopten unes estratègies de supervivència semblants; per exemple, el tritó meridional (Pleurodeles waltl) acostuma a passar l’estiu als fons de les esquerdes que es formen al fang dessecat del llit dels bassals. En aquesta mateixa zona, amb les característiques tempestes del final de l’estiu, força animals refugiats poden aparèixer fugaçment a la superfície en unes àrees reduïdes, cosa que ha donat lloc a les creences populars sobre les pluges de gripaus. Una espècie mediterrània, el gripau d’esperons (Pelobates cultripes), ha estat tot sovint protagonista d’aquestes suposades pluges, perquè de vegades apareix bruscament, en grups de milions d’individus, en una zona humitejada per una tempesta d’estiu.
Durant l’estiu, els rèptils són molt importants en les biocenosis mediterrànies. Encara disposen de prou entomofauna per a nodrir-se i llurs necessitats tèrmiques es troben cobertes. La seva reproducció és tardana: la majoria de les espècies fresen al final de la primavera o al començament de l’estiu, cosa que els permet, de vegades, una segona fresa ben entrada la tardor. Romanen actius durant tot l’estiu i es refugien si cal sols durant les hores de més calor. Com que la competència per l’aliment és intensa en l’època estival, diverses espècies han recorregut a l’especialització alimentària per tal d’alleujar-la. Per exemple, en cada regió mediterrània hi ha una o diverses espècies de petits llangardaixos mirmecòfags, que disposen de l’abundant recurs de les formigues granívores (per exemple, Blanus cinereus a la conca mediterrània, o Phrynosoma coronatum a Califòrnia). Les serps s’enfronten a una disminució de les poblacions de petits mamífers i de cries d’ocells, per la qual cosa poden adoptar estratègies de canvi de dieta, tot menjant petits lacèrtids, cargols i grans insectes, però es mantenen actives durant l’estiu.
Els ocells insectívors formen ara un percentatge menor de la població ornítica, on els ocells frugívors i granívors tenen un pes major. Per exemple, els coloms i les tórtores es reprodueixen tardanament, a la primeria de l’estiu, quan els fruits i les llavors abunden, però, per a aquestes, com per a tots els animals, la sequera estival obliga a intensificar la recerca de menjar i d’aigua. La concentració d’animals al voltant de les basses de reg o les fonts permanents en facilita molt la caça i representa un greu risc demogràfic. Els ocells rapinyaires també modifiquen la dieta i passen a dependre sobretot dels rèptils, mentre que a la primavera caçaven especialment petits mamífers. Aquests cada vegada són més escassos, fins al punt que la seva població assoleix un mínim a la tardor, cosa que obliga també els petits carnívors mediterranis a canviar de dieta: fins i tot les espècies més carnívores, com els gats mesquers, es veuen obligades a ingerir una part important de fruits en llur dieta tardorenca o de final d’estiu. Evidentment, els ocells rapinyaires poden recórrer, i ho fan molt sovint, a alimentar-se dels cadàvers d’animals que els automòbils deixen a les carreteres. En general, els vertebrats mediterranis, tret dels rèptils, disminueixen o arriben a interrompre completament llur activitat durant la canícula.
L’activitat autumnal
El màxim anyal de pluges de tardor s’inicia a la Mediterrània, amb tempestes fortes, en ple estiu. A l’inici de la tardor, l’aire calent sobre el mar afavorirà un règim pluvial típic d’aquesta zona i caracteritzat per la intensitat, la brevetat i la irregularitat de les precipitacions. Si bé la major part dels organismes ja ha acabat llur cicle anual, la nova disponibilitat d’aigua permet de rebrotar a moltes plantes i a força animals de sortir de l’estivació.
Per a alguns animals, com ara els petits rèptils, la reactivació tardorenca del medi és una oportunitat per a repetir l’esforç reproductor i per a augmentar llur població. Per a aquests i per a altres vertebrats, es tracta, fet i fet, de l’època adient per a acumular reserves d’aliment i preparar-se per al fred hivernal o, en el cas dels ocells, per a la migració. L’activitat en aquesta època no sembla dependre tant dels recursos disponibles com de ritmes endògens, sincronitzats per la disminució del nombre diari d’hores de llum i per la situació hormonal de l’organisme. Quan hi ha prou reserves acumulades, els animals acostumen a refugiar-se per passar l’hivern inactius, fins i tot si la temperatura és elevada i l’aliment abundant. Hom ha comprovat que els rèptils mediterranis, per exemple, disminueixen l’activitat i arriben a una veritable hivernació amb temperatures notablement més altes que les que suporten a la primavera, quan inicien la seva activitat reproductora.
L’activitat hivernal
L’alleujament que representen les pluges de tardor per als animals hivernants queda interromput quan arriben els primers freds. El temps encara és bo a les zones mediterrànies els primers dies d’octubre, mentre que a les terres boreals ja han caigut les primeres nevades intenses, cosa que n’ha expulsat els últims ocells migradors. El fet que aquests ocells recalin a la Mediterrània o a Califòrnia per quedar-s’hi o prosseguir llur viatge cap al sud és el senyal que s’inicia l’època hivernal per a la fauna sedentària.
Com cal esperar, són els animals heteroterms els més sensibles a les baixes temperatures, i són els que hivernen obligadament, sobretot si assoleixen unes certes dimensions. D’aquesta manera, les serps grans acostumen a hivernar, mentre que les petites, com els llangardaixos, presenten una diapausa atenuada i es refugien en caus superficials dels quals emergeixen els dies assolellats i càlids, fins i tot en ple hivern. Els insectes adults presenten una conducta similar, si bé les seves poblacions es troben molt disminuïdes, ja que acostumen a passar l’hivern en fase d’ou o en altres fases de resistència. En canvi, per als amfibis l’hivern és una estació benigna, excepte en els dominis submediterranis situats a la muntanya o en latituds altes. Així, determinades espècies modifiquen el cicle biològic anual i es reprodueixen en ple hivern en lloc de fer-ho a la primavera, com és normal en aquest grup. Aquesta circumstància es dóna no solament en representants de famílies típicament mediterrànies, com ara els discoglòssids, sinó també en espècies de latituds més altes, que a la conca mediterrània assoleixen el límit meridional de llur distribució. Per exemple, la granota roja (Rana temporaria), l’amfibi més septentrional d’Euràsia, que fresa durant la primavera i l’estiu a la major part de la seva àrea, ho fa al desembre en alguns indrets de la Península Ibèrica.