Flux d’energia i cicles de la matèria
El món que ens envolta se’ns obre davant els ulls com una realitat que es mou i es transforma contínuament. La posició dels cossos en l’espai, les formes, els colors, els sons, les olors, la temperatura, tot canvia contínuament. És un espectacle que sempre ha fascinat els humans i que, al mateix temps, els ha empès a intentar comprendre què passava al seu voltant per tal d’adaptar-se més bé a les condicions externes. Així, des de temps immemorables (probablement des que existeix), l’espècie humana s’ha anat construint esquemes mentals, models que li permetessin posar una mica d’ordre al desordre aparent de les coses, per poder preveure els canvis successius i modificar-los en benefici propi.
La definició d’ecosistema
Actualment, la ciència que estudia el medi natural, l’ecologia, ha elaborat un model força complex. Tanmateix, però, encara es basa en el concepte relativament simple de sistema: conjunt de parts interrelacionades entre si de tal manera que, si alguna d’aquestes parts canvia, afectarà les altres, i viceversa. La idea de sistema és particularment útil i s’utilitza per a descriure les relacions que s’estableixen en el nostre planeta entre el món viu i l’inert. Els sistemes formats per un conjunt d’éssers vius en contacte amb la natura inerta que els envolta són anomenats ecosistemes; un bosc, amb els seus animals, les seves plantes, el substrat on apareix, l’aigua que hi circula i la llum del sol que hi arriba, és un ecosistema, com també ho és un llac, un prat o un camp de blat.
En tots els ecosistemes hi ha un component anomenat abiòtic (del grec "abios", sense vida) i un component biològic o biòtic (del grec "bios", viu), que inclou tots els organismes que viuen en l’ecosistema considerat. Els factors abiòtics que formen part d’un ecosistema són essencialment el sòl, l’aigua i el clima, els quals, però, estan influïts profundament pel sistema viu. El conjunt dels organismes que viuen en un determinat ecosistema, agrupats en poblacions de la mateixa espècie, s’anomena comunitat (o biocenosi) del sistema. La massa global dels organismes presents en l’ecosistema en constitueix la biomassa, que es mesura en pes sec o bé en contingut d’energia, ja que la major part del pes sec és composta per carboni i hidrogen, i en els enllaços C-H hi ha continguda l’energia química dels éssers vius.
L’ecosfera. Fluxos i cicles
Els ecosistemes són realitats diferenciades, però no aïllades de la resta de la natura, i les relacions més o menys estretes que estableixen entre ells els converteixen en subsistemes d’un únic i gran sistema que engloba tot el planeta i totes les formes de vida terrestre. Aquest gran sistema rep el nom d’ecosistema terrestre, ecosfera o biosfera. Això constitueix el model global elaborat per l’ecologia per interpretar el món natural de què hem parlat al començament. Així, doncs, segons aquest model, tant la matèria viva com la inerta que constitueixen la superfície del planeta Terra formen part de l’ecosistema terrestre, però després es distribueixen pels innombrables ecosistemes parcials que componen la mateixa ecosfera (els diversos llocs on es troben els ecosistemes són anomenats biòtops).
Intentem ara mirar com "funciona" la biosfera, és a dir, observem a grans trets com es modifiquen amb el temps i com s’influeixen recíprocament les innombrables "parts" o "aspectes" que la componen.
En primer lloc observem que tots els canvis que es produeixen en la biosfera, per més variats i diferents que siguin, sempre es poden reduir a dues formes fonamentals: el flux i el cicle. Es parla de "cicle" quan una realitat física pateix transformacions que repetidament, tard o d’hora, la retornen a la forma original. En canvi, es parla de "flux" quan aquesta realitat física pateix canvis irreversibles. Hi ha fluxos i cicles que afecten la globalitat de l’ecosfera, mentre que n’hi ha d’altres que es refereixen d’una manera específica als ecosistemes individuals.
El flux de l’energia solar i el cicle de l’aigua
Un flux que afecta l’ecosfera sencera és el de l’energia irradiada pel Sol, que mesurada en termes de calor equival cada any a l’energia continguda en 1014 tones (cent bilions de tones) de carbó. Aquesta quantitat enorme d’energia és la causa de la majoria dels canvis, moviments i transformacions a què al·ludíem al començament d’aquest article i, que concerneix la matèria distribuïda per la superfície de la Terra. Una de les transformacions més espectaculars és el cicle de l’aigua, que es pot dir que es produeix contínuament sota la nostra mirada. Principalment es deu a la calor solar que evapora de la superfície de les masses líquides dels oceans una quantitat enorme d’aigua, la qual, en condensar-se a les capes altes (i fredes) de l’atmosfera, torna a precipitar sobre la superfície terrestre en forma de pluja, neu i pedra; és així com torna a la massa oceànica: o per caiguda directa o, indirectament, precipitant primer sobre la terra ferma i fluint després en forma de rius, torrents i cursos d’aigua subterranis fins a arribar a l’oceà. Es calcula que l’aigua dels oceans i de les mars constitueix el 97% de l’aigua del planeta; el 3% restant, que tanmateix és una massa considerable que equival a 47 000 km3 d’aigua, es distribueix pels rius, els llacs, el sòl i el subsòl, les glaceres i l’atmosfera (vegeu també "L’aigua com a recurs").
L’energia irradiada pel Sol arriba a tota l’ecosfera i després es distribueix, seguint modalitats diverses, per cadascun dels ecosistemes individuals. Encara més, segons els estudiosos, són precisament aquestes diverses modalitats allò que confereix una estructura diferent a cada ecosistema.
Els cicles biogeoquímics
Entre els cicles de la matèria activats per l’energia solar, són de gran importància els anomenats cicles biogeoquímics, en el curs dels quals la matèria passa contínuament de la part inanimada de l’ecosistema (abiòtica) a l’animada (biòtica) i viceversa. Els elements químics que intervenen en els cicles biogeoquímics són els mateixos que constitueixen les molècules de les cèl·lules dels organismes vius, i per això reben el nom de bioelements. Els principals, presents en tots els organismes, són el carboni, l’oxigen, l’hidrogen, el nitrogen i el fòsfor (vegeu "Biomassa i cicles de la matèria", i "Breu història del sistema viu"). Normalment els cicles biogeoquímics no afecten tota l’ecosfera sinó els diferents ecosistemes locals. Però cal tenir present que el cicle d’alguns elements químics —per exemple, el del nitrogen— no és únicament local sinó que passa per etapes que actuen sobre tot l’ecosistema del planeta. De fet, tots els subecosistemes capten nitrogen de la "reserva" atmosfèrica, i la concentració de nitrogen lliure és la mateixa a tota l’atmosfera. L’obtenció de nitrogen de l’atmosfera, i el seu retorn, no és a càrrec de cada ecosistema parcial sinó, globalment, de tot el sistema viu, o bé de tota la biocenosi terrestre (vegeu, més endavant, "Biomassa i cicles de la matèria").
La vida, el clima i el sòl
Més amunt ja hem dit que a l’ecosfera, com a sistema, les diferents parts que la componen es modifiquen al llarg del temps i s’influeixen recíprocament. Vegem ara alguns exemples importants d’aquesta interdependència, com ara la influència dels factors abiòtics sobre el clima, la influència del sòl sobre la vida i la manera com la vida modifica el sòl i influeix sobre el cicle de l’aigua i sobre la llum.
Abiosi, fotosíntesi i clima
L’exemple més significatiu que podem considerar és potser el del clima, pel que fa als aspectes bàsics com la temperatura, la humitat, les precipitacions i les seves variacions periòdiques. En part, les diferències climàtiques que trobem entre les diverses regions del planeta depenen de condicions abiòtiques. En primer lloc depenen de la latitud, és a dir, de la distància de l’equador, amb una temperatura mitjana que serà més alta com més ens hi acostem. L’amplitud de les oscil·lacions tèrmiques estacionals també depèn de la distància de l’equador, mentre que la variació de temperatura que es produeix al llarg de les 24 hores (és a dir, la variació tèrmica diària) es veu afectada a més per altres factors abiòtics com l’altitud i, per tant, la configuració del relleu (les muntanyes, els vessants, etc.) i la naturalesa del sòl. La proximitat de la mar, o dels grans llacs, també influeix en l’oscil·lació tèrmica, que no és tan brusca. Amb certa periodicitat es verifiquen les glaciacions, fenòmens que poden ser atribuïts a esdeveniments geològics o astronòmics.
Els volcans tenen, així mateix, certa influència sobre el clima, ja que durant les erupcions emeten cendres i pols que intercepten parcialment la llum solar i provoquen un descens de la temperatura. D’altra banda, els volcans també han produït i produeixen encara l’efecte contrari d’augmentar la temperatura a través de l’emissió de diòxid de carboni; és el que s’ha anomenat "efecte hivernacle", del qual es parla més àmpliament a "Els danys mediambientals del desenvolupament".
Fins ara, hem considerat els components abiòtics més importants que influeixen sobre la determinació del clima. Però la comunitat viva també hi influeix a través de les funcions fonamentals: la fotosíntesi i la respiració.
La fotosíntesi que du a terme la comunitat vegetal (com s’il·lustra a "El món dels vegetals") transforma el carboni inorgànic en orgànic, en molècules de glucosa; si es considera el fenomen per a cada àtom de carboni implicat, es verifica que una molècula de diòxid de carboni, CO2, es combina amb una molècula d’aigua, H2O, i dóna lloc a la formació del grup CH2O (constitutiu de la molècula de glucosa) i a l’alliberament d’una molècula d’oxigen, O2. En la respiració, la reacció procedeix en sentit invers: combina un grup de CH2O amb una molècula de O2 i torna a obtenir els materials de partida, una molècula de CO2 i una molècula de H2O (vegeu "La cèl·lula").
Durant molt de temps la biocenosi terrestre ha realitzat molt més la fotosíntesi que la respiració, és a dir, ha obtingut amb la primera operació més CO2 del que ha alliberat amb la segona. D’aquesta manera ha impedit (o ha contribuït a impedir) que l’emissió contínua de CO2 dels volcans conduís a un augment de la seva concentració a l’atmosfera i, per tant, a un desenvolupament de l’efecte hivernacle, amb el consegüent augment de la temperatura. Així, la superioritat de la fotosíntesi respecte de la respiració ha contribuït a estabilitzar el clima (vegeu també "Breu història del sistema viu").
La part del material sintetitzat per la fotosíntesi que no ha estat sotmesa a la respiració ha patit transformacions químiques al llarg del temps que l’han feta inservible per a alimentar tots els organismes vius; aquest és l’origen dels jaciments de combustibles fòssils (petroli, metà i carbó).
El predomini de la fotosíntesi sobre la respiració, que era mínim al principi de la vida de les plantes (fa potser 3 500 milions d’anys) i que després va augmentar a gran velocitat, va perdurar fins al Carbonífer (entre 345 i 270 milions d’anys). En aquella època, la fotosíntesi i la respiració es van començar a anivellar i es va iniciar una fase d’equilibri que encara perdura. Probablement podem situar en aquell període el començament de l’ecosistema terrestre del qual formem part i que, des de fa aproximadament 10 000 anys, els humans hem començat a modificar.
És possible que les mateixes plantes regulin l’oxigen, mitjançant la reducció de la fotosíntesi si la seva concentració augmenta.
També és possible, però, una relació entre factors biològics i factors abiòtics, perquè cada vegada que la concentració d’oxigen atmosfèric augmenta provoca un increment dels incendis espontanis o un augment de la gravetat dels incendis provocats i, per tant, a través dels processos de combustió, una disminució de l’oxigen atmosfèric i una emissió de diòxid de carboni més gran.
La teoria de Gaia
La temperatura, la qualitat del sòl, el cicle de l’aigua i la qualitat de la llum són condicions aparentment abiòtiques de l’ecosistema terrestre (i en alguns casos, com el cicle de l’aigua o la qualitat del sòl, també caracteritzen els subsistemes concrets). Només un estudi una mica aprofundit ens revela que aquestes condicions aparentment abiòtiques són en realitat el resultat de complexos intercanvis entre els components abiòtics i la comunitat viva. L’observació d’aquests fenòmens porta alguns científics a considerar el planeta com un únic organisme viu, capaç d’autoregular-se per tal de superar els factors adversos que posarien en perill la supervivència de la biocenosi, i donen a aquesta interpretació el nom d’"hipòtesi de Gaia" (del mot grec que designa la Terra i del qual provenen els termes geografia, geologia, etc.). El científic britànic James Lovelock, el primer teòric de "Gaia", opina que "el nivell actual d’oxigen es troba en un punt en què els riscs i els beneficis s’equilibren de manera adequada"; ara bé, cal observar que aquesta valoració d’adequació no es refereix a les exigències de tota la comunitat viva, sinó a les exigències de tan sols una part, la part de la biocenosi que no solament és capaç de tolerar l’oxigen (els organismes aerobis) sinó que no podria sobreviure en un medi no oxigenat. No es podria fer la mateixa valoració si es considerés la part de la comunitat que ha mort perquè no ha aconseguit tolerar l’oxigen o la part de la comunitat que només pot viure en medis especials mancats d’oxigen (anaerobis) com els fons de les aigües eutrofitzades. Per què s’escull un criteri de valoració que, per dir-ho d’alguna manera, adopta el punt de vista dels organismes aerobis i no el dels organismes anaerobis? Per a respondre aquesta pregunta s’han de considerar dues possibilitats.
La primera possibilitat és que aquesta elecció s’ha fet perquè els organismes aerobis constitueixen la gran majoria de la comunitat viva. Però aquest fenomen depèn de l’adaptació: en un medi oxigenat, el fet que la major part de la comunitat sigui aeròbia no revela tant una millor predisposició a viure general com, simplement, una millor predisposició a viure en aquestes condicions. I el fet que els organismes aerobis, poquíssims al començament, hagin crescut desmesuradament pot explicar-se per la llei formulada per l’economista i demògraf anglès Thomas Robert Malthus al començament del segle XIX sobre la gran quantitat de naixements enfront de les possibilitats de sobreviure: un malbaratament de naixements (de vides, d’energia) que té com a conseqüència, d’una banda, el creixement de la biomassa fins a assolir una situació d’estabilitat o de clímax i, de l’altra, l’evolució del sistema viu a través de la selecció, de manera que qualsevol canvi de les condicions abiòtiques de l’ecosistema genera processos d’adaptació de la comunitat viva que premia els organismes més adaptables a les noves condicions.
L’altra possibilitat és que darrere l’elecció d’adoptar el punt de vista dels aerobis hi hagi el prejudici d’assignar a aquests organismes més importància que als anaerobis. Darrere aquest prejudici hi ha, més o menys conscient, la interpretació que l’evolució biològica ha tingut com a finalitat l’aparició de l’espècie humana, que en seria la culminació. Es tracta d’una idea antiquada que hem d’abandonar per tal d’entendre el funcionament de l’ecosistema i per saber-lo respectar, així com per garantir-ne la supervivència, amb la qual cosa també en garantim la nostra. De fet, encara és molt estesa (per bé que hagi començat a qüestionar-se) la convicció que l’espècie humana pot adoptar davant la natura una actitud de domini i d’explotació, i darrere aquesta convicció hi ha la seguretat que totes les nostres exigències seran satisfetes perquè l’espècie humana és el significat i la culminació del llarg camí de la vida. Aquesta seguretat posa en perill el conjunt de l’ecosistema i, per tant, també a nosaltres, que en formem part.
La influència del sòl sobre la vida
El fet que, l’ecosistema terrestre hagi començat a estabilitzar-se entre 400 i 300 milions d’anys enrere, i que encara es mantingui estable, no vol dir que des d’aleshores no hagi patit cap canvi. En efecte, aquesta estabilització només afecta l’equilibri global d’alguns aspectes de l’ecosfera, però la biosfera és sotmesa a canvis continus, i en l’àmbit local, en els ecosistemes concrets, els canvis són gairebé incessants. Quan un ecosistema troba una certa estabilitat es diu que ha assolit el "clímax", i la seva comunitat viva s’anomena "comunitat clímax".
La composició d’una comunitat clímax depèn en bona part del clima. Tanmateix, en una regió climàticament homogènia hi pot haver associacions vegetals diferents, és a dir, comunitats diferents (normalment, d’una comunitat tan sols es consideren les espècies vegetals perquè, en ser fixades al sòl, són seleccionades per les condicions ambientals més clarament que els animals, els quals es poden desplaçar i tenen més possibilitats d’evitar les seleccions més severes). L’existència de comunitats diverses en una regió climàticament homogènia depèn de les variacions de les característiques del sòl. Normalment, pel que fa a les regions on s’observa un clima homogeni, són més àmplies aquelles que es caracteritzen per l’homogeneïtat del sòl, és per això que es distingeix el clímax climàtic dels clímaxs edàfics (del mot grec "edaphos", sòl).
La millor manera de comprendre aquests conceptes és recórrer a un exemple: considerarem les associacions vegetals d’una regió de l’Ontario meridional (Canadà), homogènies des del punt de vista climàtic. La regió és en la seva extensió màxima gairebé plana i suaument ondulada, amb alguns turons; es parla de "microclima normal" en la zona plana, de "microclima fred" en els pendents septentrionals i de "microclima càlid" en els pendents meridionals. Hi ha zones de sòl humit, d’altres de sòl eixut i d’altres de sòl mal drenat (pantanós). Les condicions climàtiques i les edàfiques formen, doncs, noves combinacions diferents, amb noves associacions diferents d’arbres. Però la situació no és estàtica: l’alzina i la cària (un tipus de noguera) viuen a la plana sobre sòl eixut; la pèrdua anual de fulles i branques, i les seves despulles després de la mort, proporcionen al sòl la matèria orgànica que reté l’aigua (vegeu, més endavant, "La vida, el cicle de l’aigua i la llum"); el sòl eixut es modifica, esdevé més humit, i l’associació d’alzina i de cària és substituïda progressivament pels aurons i els freixes. Per un procés anàleg, en els pendents encarats al sud l’associació de cària i castanyer es transformarà a causa de la substitució del castanyer per la magnòlia; en els pendents exposats al nord, l’avet de Douglas i el bedoll seran substituïts pels oms, els freixes i les alzines.
La vida, el cicle de l’aigua i la llum
Si el clima, com hem vist, és àmpliament influït per la comunitat viva, aquesta, al seu torn, també modifica el sòl (com, d’altra banda, ja hem dit en el paràgraf anterior). En primer lloc, les arrels de les plantes penetren en els intersticis de la roca de sota i en créixer de diàmetre els eixamplen, fins que la roca s’esquerda. L’acció es prolonga fins després de la mort de la planta, perquè en els intersticis eixamplats i després buidats per la descomposició de les arrels s’insinua l’aigua, que durant l’hivern es glaça i per tant es dilata i fa pressió contra la roca fins que la trenca. A més d’aquesta acció mecànica, també hi ha una acció de caràcter químic causada per substàncies dissoltes en l’aigua (com el diòxid de carboni, que es transforma en àcid carbònic) o secretades per les arrels, que actuen sobre les partícules de roca com més fragmentades estan (grava, sorra, llim), i les modifiquen. Així es formen els minerals argilosos.
La comunitat viva cedeix al sòl la matèria orgànica (molècules construïdes amb els productes de la fotosíntesi) que serveix d’aliment als nombrosos microorganismes que conté. Els aerobis (és a dir, els que necessiten oxigen per a sobreviure), mitjançant la respiració, restitueixen al medi el diòxid de carboni i l’aigua; però al sòl també hi ha molts microorganismes anaerobis, els residus dels quals encara contenen carboni i hidrogen. Els microorganismes del sòl, les seves despulles, els seus residus, les substàncies químiques que produeixen formen l’humus, una substància que cobreix les minúscules partícules sòlides del sòl, que les agrega i obstaculitza la pèrdua lenta de l’aigua, i reté les sals minerals. Quan un sòl conté una mescla ben proporcionada de sorra fina, argila i humus, es revela molt adequat per al creixement de les plantes: el component sorrenc el fa permeable a l’aire i per tant les arrels poden respirar, l’argila i l’humus retenen aigua i sals, i les molècules orgàniques produïdes pels microorganismes proporcionen substàncies útils a les plantes que fan funcions semblants a les de les vitamines. Així, les plantes no solament participen en la configuració del clima, sinó també en la formació del sòl.
Com hem vist més amunt, la comunitat vegetal influeix sobre el cicle de l’aigua en proporcionar al sòl la matèria orgànica destinada a transformar-se en humus. D’aquesta manera alenteix el ràpid escorriment de les aigües superficials (que arrossegaria el sòl i deixaria les roques al descobert) i s’assegura l’aigua en èpoques de sequera. Però les plantes també interfereixen en el cicle de l’aigua d’altres maneres: en efecte, un arbre funciona com una bomba, que xucla l’aigua de les profunditats del sòl a través de les arrels i la transporta fins a les fulles, on s’evapora; igualment, també pot absorbir la humitat de l’aire i l’aigua de pluja.
La biocenosi també actua sobre la llum, ja que, en alliberar oxigen, la gran quantitat de O2 acumulada en l’aire per la comunitat viva ha originat al voltant del planeta la formació d’una capa d’ozó que el protegeix de les freqüències més perilloses de les radiacions ultraviolades (vegeu "Breu història del sistema viu"). El component ultraviolat que travessa la capa d’ozó i arriba a la comunitat viva és certament indispensable, sobretot per als animals que el necessiten per a la fixació del calci. Els excessos són perjudicials: es considera que un excés d’ultraviolats és una de les causes, per exemple, del creixement escàs de les plantes per sobre d’una certa altitud.
Biomassa i cicles de la matèria
La biomassa, és a dir, la massa dels organismes vius, és formada per matèria orgànica, caracteritzada per molècules que contenen carboni i hidrogen. És la mateixa composició de la necromassa (és a dir, la massa dels òrgans vegetals morts però que encara no s’han desprès de les plantes vives), la virosta (les parts mortes caigudes al sòl) i l’humus, que s’origina a partir de les transformacions de la virosta i la descomposició dels residus animals. Dels 92 elements químics existents en la natura, la matèria orgànica en conté aproximadament una quarantena. Són els bioelements als quals ja hem al·ludit; d’aquests, només cinc són indispensables per a tots els organismes: C (carboni), H (hidrogen), O (oxigen), N (nitrogen) i P (fòsfor). Aquests cinc elements, doncs, es distribueixen en diferent concentració per tota la matèria orgànica. Hi ha altres bioelements que tan sols apareixen en una part de la matèria orgànica, la qual cosa no vol dir que siguin menys importants: el magnesi (Mg), per exemple, és indispensable per a les plantes perquè forma part de la molècula de clorofil·la, sense la qual no hi pot haver fotosíntesi. Així, malgrat no ser indispensable per a tots els organismes, el magnesi resulta imprescindible per al conjunt del sistema viu, i per a cada ecosistema concret.
Com ja sabem, els bioelements participen en cicles de matèria anomenats biogeoquímics (vegeu, més amunt, "El flux de l’energia solar i el cicle de l’aigua"). Ara ens ocuparem de dos d’aquests cicles: el del nitrogen i el del fòsfor. Pel que fa als del carboni, l’hidrogen i l’oxigen, estretament relacionats amb la història evolutiva del sistema viu, s’ha cregut oportú parlar-ne a "Breu història del sistema viu".
El cicle del nitrogen
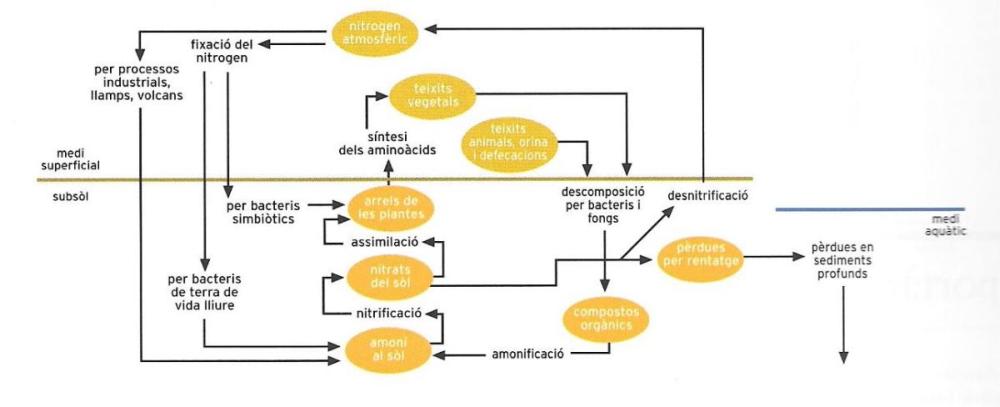
El nitrogen es desplaça de manera cíclica entre l’atmosfera, el sòl i els éssers vius. L’activitat quimiosintetitzadora dels bacteris del sòl transforma el nitrogen atmosfèric en compostos nitrogenats. Al sòl, també hi van a parar els cadàvers i els productes de rebuig, que hi aporten més nitrogen. Els bacteris nitrificants el transformen de manera que les plantes el poden absorbir i convertir en aminoàcids, amb els quals sintetitzaran les seves proteïnes; així, el nitrogen entra al sistema dels éssers vius.
ECSA
El nitrogen, com a constituent de les substàncies proteiques i dels àcids nucleics, té una importància cabdal per als éssers vius. La quantitat més gran d’aquest element es troba a l’atmosfera (aproximadament el 78% del volum) en estat molecular. Tot i això, només uns quants organismes són capaços d’utilitzar directament el nitrogen molecular o, com se sol dir, de fixar el nitrogen atmosfèric (els organismes fixadors del nitrogen), ja que la majoria dels organismes utilitzen el nitrogen només de forma combinada.
Tots els éssers vius que tenen capacitat, afortunadament per a ells (i per a nosaltres: vegeu l’insert "La importància dels procariotes"), de fixar el nitrogen atmosfèric són procariotes. Es tracta de diversos gèneres de bacteris i de les anomenades algues blaves o cianobacteris. Les plantes no són capaces de fixar el nitrogen, i els animals encara menys. Els procariotes fixadors del nitrogen viuen arreu: els cianobacteris a l’aigua i als sòls humits , i els bacteris típics preferentment al sòl. La seva manera de viure i de nodrir-se varia molt: viuen pel seu compte o bé en simbiosi amb altres organismes; n’hi ha que viuen d’oxigen (aeròbics) i altres que l’eviten (anaeròbics); n’hi ha d’autòtrofs (els cianobacteris i alguns bacteris típics) i d’heteròtrofs. La capacitat fotosintetitzadora dels autòtrofs és estretament relacionada amb la fixació del nitrogen.
La fixació del nitrogen és una mena de reacció química on el nitrogen present en l’atmosfera es combina amb l’oxigen, és a dir, es tracta d’una reacció d’oxidació. Els organismes fixadors se serveixen del nitrogen fixat per a produir les molècules orgàniques nitrogenades del mateix organisme. Alhora alliberen compostos nitrogenats, els nitrats, que es dissolen en el sòl i són absorbits per les plantes, les quals els utilitzen per a incorporar-los en forma orgànica a les seves proteïnes i als seus àcids nucleics.
La molècula de nitrogen atmosfèric (N2) és una molècula "forta", i per a trencar-la cal molta energia, la qual pot ser proporcionada pels raigs ultraviolats del Sol o per la descàrrega elèctrica dels llamps. En aquest últim cas una petita part del nitrogen atmosfèric es transforma en nitrats, que es precipiten al sòl amb la pluja. És molt probable que els organismes vius més antics, encara incapaços de fixar el nitrogen, hagin rebut per aquesta via els nitrats necessaris; però fins i tot després d’haver adquirit aquesta capacitat, la contribució de la pluja nítrica va continuar essent preciosa, perquè la fixació del nitrogen constitueix per als organismes vius una gran despesa energètica.
L’eficàcia del procés va millorar notablement després que el sistema viu s’independitzés del medi aquàtic i ocupés gradualment les terres emergides. L’evolució successiva de les plantes va portar a altres simbiosis; algunes plantes, com per exemple el vern, l’araucària i el gingko, van entrar en simbiosi amb fongs primitius, unicel·lulars, també capaços de fixar el nitrogen; les lleguminoses van entrar en simbiosi amb els bacteris del gènere Rhizobium, els quals corresponen a l’hospitalitat de les plantes fixant el nitrogen atmosfèric, cosa que també els és favorable.
D’altra banda, la simbiosi amb vista a fixar el nitrogen no tan sols ofereix avantatges als simbionts, sinó també a la vegetació dels voltants. En efecte, les despulles de les lleguminoses contenen més proteïnes que les de les altres plantes, i en descompondre’s formen un humus amb abundant nitrogen fixat. Com que la vegetació en general creix millor si està barrejada amb plantes associades a fixadors del nitrogen, els organismes heteròtrofs també en surten beneficiats. Un dels pilars de la conservació i la prosperitat del sistema viu és, en definitiva, la fixació simbiòtica del nitrogen.
Ja hem dit que els nitrats serveixen a les plantes per a produir les seves molècules orgàniques nitrogenades (les proteïnes i els àcids nucleics). Després hi ha dos camins possibles: les plantes poden servir d’aliment als animals, o bé als fongs i als bacteris descomponedors que aprofiten la necromassa (fulles mortes, branques seques, etc.) que s’ha separat de la planta i les plantes senceres que han caigut en morir. En aquest segon cas, la descomposició duta a terme pels bacteris i els fongs dóna lloc a la formació d’un altre compost nitrogenat, l’amoníac, la molècula del qual, NH3, és constituïda per nitrogen i hidrogen.
L’amoníac, com els nitrats, pot ser utilitzat per les plantes o bé pot ser oxidat i transformat en nitrit (que al seu torn s’oxida en nitrat). El nitrogen, doncs, es troba altre cop amb dues possibilitats, ja que els nitrats són utilitzats per les plantes o bé són sotmesos a l’acció de bacteris que el redueixen a N2 i el retornen a la "reserva" atmosfèrica.
En canvi, si les plantes són menjades pels animals, el nitrogen, després d’haver romàs a l’interior dels seus cossos (dels quals constitueix bona part de la biomassa com a component de les proteïnes i els àcids nucleics), és expulsat amb les dejeccions (la urea i l’àcid úric per l’orina, les proteïnes per la femta) o, un cop morts, amb les despulles; i així, com el nitrogen contingut en els residus i les despulles dels microorganismes fixadors del nitrogen i de les plantes, també reprèn el procés que el redueix a amoníac.
Fem ara un breu resum del cicle del nitrogen. És un laberint molt complicat on el nitrogen es presenta de quatre formes diferents: nitrogen lliure (N2), nitrogen nítric (oxidat), nitrogen amoniacal (reduït) i nitrogen orgànic (en els cossos dels organismes vius i en els seus residus). El nitrogen s’introdueix en els organismes vius per un procés d’oxidació i hi circula pels diferents "compartiments", els abandona per un procés de reducció que el transforma en nitrogen amoniacal, i es torna a incorporar al sistema viu, que el retorna a l’atmosfera. Aquesta última fase s’anomena desnitrificació, i els bacteris que en són responsables, desnitrificadors.
La reserva atmosfèrica de nitrogen té, doncs, dues portes que donen a la biosfera, una que li cedeix nitrogen (la fixació del nitrogen) i l’altra que el torna a acollir (la desnitrificació). Totes dues portes són "gestionades" per organismes unicel·lulars primitius: fongs o bacteris.
La importància dels procariotes
La capacitat d’utilitzar directament el nitrogen atmosfèric (és a dir, de fixar-lo) és pròpia només de diversos organismes procariotes. El fet que els únics organismes capaços de portar a terme una funció tan fonamental per a la vida de tots els éssers vius siguin els modestos procariotes ens porta a considerar aquests éssers minúsculs amb més respecte del que nosaltres, els humans, des de la nostra posició d’organismes complexos i evolucionats, estem disposats a atorgar-los. Però la nostra actitud de suficiència hauria de canviar encara més radicalment si és certa (com sembla que ho és) la teoria segons la qual, en darrer terme, la capacitat de les plantes de fixar l’energia solar també depèn d’organismes procariotes o dels seus descendents.
La teoria a què al·ludim defensa que els orgànuls de les cèl·lules vegetals que realitzen la fotosíntesi, els cloroplasts, no són altra cosa que procariotes que van establir-se en les cèl·lules com a simbionts i que després s’han transformat en part integrant seva. Així, doncs, la preuada fotosíntesi seria obra exclusiva o de procariotes actuals, els cianobacteris, o d’"exprocariotes", els cloroplasts de les plantes. I també els mitocondris, que són els orgànuls de les cèl·lules de tots els eucariotes destinats a la respiració, haurien tingut el mateix origen. Com a prova d’això s’addueixen diversos fets. Tant els cloroplasts com els mitocondris tenen DNA propi, diferent del de la cèl·lula hoste; són capaços d’autoreplicar-se a l’interior de la cèl·lula; s’ocupen de manera autònoma de la síntesi d’un cert nombre de proteïnes, segons modalitats que s’assemblen molt als processos anàlegs dels bacteris procariotes, i són recoberts d’una membrana doble, la més externa de les quals recorda el plasmalemma (la membrana que envolta el protoplasma) dels eucariotes, mentre que la més interna (per l’absència de colesterol) s’assembla molt més al plasmalemma dels procariotes.
El camí del fòsfor. Flux o cicle?
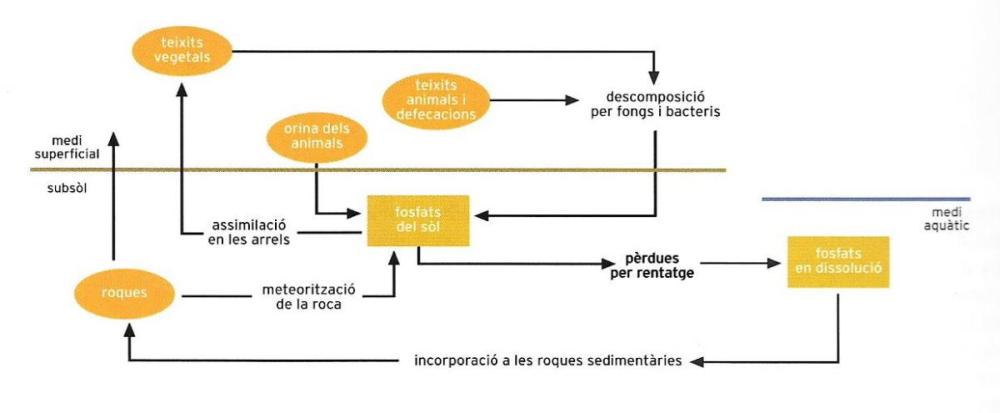
L’aigua solubilitza el fòsfor dels jaciments minerals i el posa a disposició de les plantes, que l’absorbeixen a través de les arrels i l’incorporen a la seva matèria orgànica. D’allà passarà als animals, on, juntament amb el calci, acomplirà una important missió en la formació dels esquelets. Quan les plantes i els animals moren, el fòrfor retorna al sòl. Una part, però, va a parar primer a la mar, i s’incorpora als peixos i als ocells aquàtics.
ECSA
El fòsfor (P) és necessari per a tots els organismes vius sense excepció, per a dues funcions fonamentals. Una és la funció energètica, ja que el fòsfor és contingut en la molècula d’ATP (trifosfat d’adenosina), l’única que, a l’interior de la cèl·lula viva, pot portar l’energia al punt on s’ha d’utilitzar i deixar-la disponible per al seu ús (quant a la funció de l’ATP, vegeu "La cèl·lula"). La segona funció fonamental del fòsfor és la constitució dels grups fosfat dels àcids nucleics.
En comparació amb els altres quatre elements indispensables per a la vida, el fòsfor presenta una particularitat: tan sols pot ser reciclat localment, sense participar en un sol cicle global. Pel que fa al carboni, l’aire i l’aigua tenen un dipòsit de diòxid de carboni que la comunitat viva utilitza per a la fotosíntesi i que allibera amb la respiració, de la mateixa manera que hi ha un dipòsit d’oxigen que la biocenosi empra per a la respiració i que allibera amb la fotosíntesi; i, tal com hem vist una mica més amunt, en l’atmosfera també hi ha un dipòsit de nitrogen que la biocenosi fa servir per a fixar el nitrogen i que allibera amb la desnitrificació. Posem un exemple d’ecosistema limitat: en els processos de descomposició, els residus orgànics d’un estany originen la formació de compostos del fòsfor, els fosfats, que es tornen a introduir dins els organismes vius i recorren la cadena alimentària (vegeu, més endavant, "La relació alimentària i l’especialització") fins a la descomposició següent.
D’altra banda, si considerem el camí del fòsfor des del punt de vista de l’ecosistema terrestre, podem veure que hi ha dos dipòsits de fosfats: un al començament del camí i l’altre al final, sense possibilitat que el segon retorni al primer. El dipòsit inicial dels fosfats es troba en les roques: un cop les roques han estat erosionades, les aigües arrosseguen els fosfats als rius, als llacs, als pantans, a la mar, és a dir, als llocs on la vida prospera amb més força i des d’on, transportats principalment pels animals, són escampats per les zones dels voltants. Són sobretot els ocells els que s’encarreguen d’aquesta important funció ecològica, quan es nodreixen de peixos i altres animals aquàtics i després abandonen les seves dejeccions pel sòl dels voltants, on es formen dipòsits de guano. Però així no s’aconsegueix el reciclatge total, ja que, en passar de la fase abiòtica a la biològica, el fòsfor té dues possibilitats de reciclatge parcial: una que el reté en la fase biològica a través de la cadena alimentària i una altra que, a través de l’acció dels ocells (els dipòsits de guano), el retorna a la fase abiòtica. El reciclatge és parcial, com acabem de dir, perquè una part dels fosfats dissolts en les aigües es diposita al fons dels oceans. Així doncs, el fòsfor experimenta més aviat un flux que no pas un cicle, si bé és un flux amb cicles parcials.
La vida i els seus problemes
Des de la seva aparició, la vida s’ha anat escampant poderosament pel nostre planeta, que s’ha anat poblant de milions d’espècies. Però no s’ha de creure que ha estat una victòria fàcil. En totes les èpoques anteriors i en l’actualitat, cada espècie ha hagut i ha d’afrontar obstacles i limitacions al seu creixement (de vegades sense reeixir), establir interaccions complexes (de competència i/o de cooperació) amb les altres espècies i adaptar-se a les fèrries lleis que determinen la formació i l’evolució dels ecosistemes al llarg del temps. Ara ens centrarem en els aspectes més importants d’aquest exercici de supervivència.
El factor limitant
El fet que el fòsfor presenti més aviat un flux que un cicle provoca a la biocenosi certes dificultats a propòsit d’aquest element, que en moltes situacions pot escassejar i esdevenir un factor limitant. Però, què entenem per factor limitant?
Posem-ne un exemple. Els ous de granota eclosionen si en l’ambient circumdant es donen les condicions que ho permetin, sobretot si la temperatura del medi no és inferior a 0°C ni superior a 12°C. La temperatura ideal és de 4°C. Com més s’acosta al límit inferior de 0°C o al superior de 12°C, més s’alenteix el fenomen. Doncs bé, en aquest cas podem considerar la temperatura com el factor limitant del naixement de les granotes. De la mateixa manera, si diem que en un ecosistema el fòsfor funciona com a factor limitant volem dir que, per a algunes espècies de l’ecosistema en qüestió, la quantitat de fòsfor present en limita el creixement potencial. Així, doncs, podem afirmar que, en general, el factor limitant de determinades espècies en un cert ecosistema és aquell factor que tendeix a limitar el seu possible creixement. També pot passar que el factor limitant baixi per sota del mínim que permet la manifestació d’un fenomen biològic determinat (una temperatura per sota dels 0°C en el cas del naixement de les granotes): en aquest cas, en què la supervivència d’una espècie és en joc, no es parla de factor limitant sinó de factor regulador. No tots els elements d’un ecosistema són factors limitants o reguladors. En efecte, n’hi ha que no tenen cap influència sobre el funcionament de l’ecosistema, encara que hi siguin presents en gran quantitat. Saber quins són els factors limitants i quins no resulta, doncs, molt útil per a orientar-se en la complexitat del món natural. Si, per exemple, en un curs d’aigua hi ha una mortaldat de peixos, l’ecòleg sap que és oportú comprovar-ne de seguida la concentració d’oxigen, ja que és, certament, un factor limitant i regulador de la vida dels peixos. Va ser el científic alemany Justus Liebig qui va formular el concepte de factor limitant. Es va fixar en el fet que l’abundància de les collites agrícoles depèn de la presència d’alguna substància que, en quantitat mínima, pot ser suficient, però que si apareix en quantitats inferiors a aquest mínim, o és del tot absent, posa en perill el desenvolupament dels conreus i, per tant, la collita final. A partir d’aquesta observació va enunciar la llei segons la qual "el creixement d’una planta depèn de la presència de l’element assimilable en menor concentració", i que és coneguda com a llei del mínim. Amb tot, l’exemple de les granotes ens mostra que la supervivència d’una espècie pot resultar limitada o compromesa no solament per un fet d’escassetat i de mínim, sinó també d’excés o de màxim: si la temperatura puja per sobre de 12°C, les granotes també deixen de néixer. Això porta a afirmar que els factors limitants fixen una àrea de tolerància dins la qual un ecosistema concret resulta habitable per a determinades espècies.
La relació alimentària i l’especialització
En descriure els cicles dels elements indispensables per a viure, sovint hem al·ludit a la relació alimentària entre els organismes vius. Per exemple, hem vist que el nitrogen passa de les plantes als animals en la mesura que els animals s’alimenten de plantes. La relació alimentària és una forma fonamental de relació entre els éssers vius, perquè és a través seu que els fluxos d’energia, com també els fluxos i els cicles de la matèria, recorren la biocenosi (vegeu també "Breu història del sistema viu"). Segons la seva manera d’alimentar-se, els éssers vius que formen part d’una biocenosi es poden agrupar en tres categories:
1. Els productors. Són els organismes autòtrofs o fotosintetitzadors (plantes o algues) capaços d’obtenir substàncies orgàniques a partir de substàncies inorgàniques (organificació del carboni: vegeu "Les plantes del nostre entorn"). Aquestes substàncies constitueixen la base alimentària dels altres dos grups.
2. Els consumidors. Són els organismes heteròtrofs, incapaços de produir substàncies orgàniques i que es nodreixen, directament o indirectament, de les elaborades pels productors. Se subdivideixen en consumidors de primaris (herbívors), consumidors secundaris (carnívors que mengen herbívors) i consumidors terciaris (carnívors que s’alimenten de carnívors que, al seu torn, es nodreixen d’herbívors). Tots els animals, inclosos nosaltres, formem part d’aquesta categoria.
3. Els descomponedors. Són els organismes que es nodreixen de la biomassa morta produïda per les dues categories anteriors. La glucosa sintetitzada per la fotosíntesi, és a dir, produïda per l’organificació del carboni, pot proporcionar energia a través d’escissions que no necessiten oxigen, com la glicòlisi, que provoca la formació d’àcid làctic en els teixits musculars, o bé les diverses formes de fermentació (vegeu "La cèl·lula"). Però la glucosa proporciona quantitats d’energia molt més grans si se sotmet a oxidació durant el procés respiratori, que restitueix al medi CO2 i H2O. Aquest procés es dóna en les tres categories esmentades.
Però analitzem amb més detall el problema de les relacions alimentàries. Les més simples són les d’una "cadena" en què cada presa té un sol predador i cada predador una sola presa, és a dir, quan totes les espècies que hi participen són especialitzades. L’exemple més conegut el trobem als llocs més àrids de la tundra dels altiplans àrtics. L’únic productor (és a dir, l’única espècie que practica la fotosíntesi) és el liquen, l’únic consumidor de primer ordre és la llebre i l’únic consumidor de segon ordre és el linx. En aquest cas l’especialització és absoluta, ja que no hi ha més espècies (llevat dels microorganismes de la descomposició), i la comunitat viva (la biocenosi) es caracteritza per una simplicitat extrema. Com més especialització hi ha i més simple és la biocenosi, més eficaç és el control que una població exerceix sobre les altres. Posem-ne un exemple: si les llebres augmenten de nombre, l’excés de pasturatge fa disminuir la biomassa dels líquens i l’abundància de preses fa augmentar la població dels linxs; però tant la disminució dels líquens com el gran nombre de linxs fan minvar la població de llebres. Això provoca un augment de la massa de líquens i una disminució del nombre de linxs, amb la qual cosa l’equilibri queda restablert. Si les condicions romanen constants s’estableix una relació quantitativa estable entre la massa de líquens, el nombre de llebres i el nombre de linxs, una mena de relació que en la realitat és poc freqüent, però que constitueix el punt al voltant del qual la relació oscil·la contínuament (en realitat el liquen té cicles d’onze anys, probablement relacionats amb fenòmens solars, i per tant les poblacions de llebres i de linxs també tenen cicles semblants, per bé que lleugerament endarrerits respecte dels del liquen).
Vegem ara quins poden ser els desavantatges d’una especialització excessiva. Per entendre-ho millor posarem alguns exemples: si un territori té molts cérvols i altres herbívors de talla mitjana o gran, el llop, tot i que no és "especialitzat", prefereix caçar cérvols i altres herbívors, però si no troba aquests animals s’adapta a l’alimentació a base de ratolins. En canvi, la vida d’una espècie molt especialitzada corre més riscs: moltes larves de papallones es nodreixen sovint d’una sola espècie vegetal: si la població de què s’alimenten aquestes larves disminueix notablement (per malaltia o per condicions climàtiques adverses), en aquell territori la població de les papallones pot córrer el perill d’extingir-se. Per contra, si una espècie com el llop pot caçar més d’una espècie, és menys probable que la seva població pugui desaparèixer.
Si una biocenosi es caracteritza per la presència de poblacions poc especialitzades, té una estructura alimentària "de xarxa", més complexa però més estable que una sèrie "de cadena". La complexitat d’un ecosistema depèn del nombre d’interaccions entre les espècies que caracteritzen la biocenosi. Generalment, un ecosistema complex és més estable que un de simple. La diversitat també és un factor d’estabilitat i es distingeix de la complexitat: per exemple, dos boscos poden ser igualment complexos, si són formats per les mateixes espècies, i per tant tenen el mateix nombre d’interaccions entre elles, però en un hi pot haver una població d’arbres de la mateixa edat, i en l’altre, la població de la mateixa espècie amb exemplars d’edats diferents. En aquest segon cas l’estabilitat és més gran, ja que la capacitat de defensar-se dels organismes patògens varia amb l’edat.
Les interaccions entre espècies poden afectar diverses espècies de plantes, una planta i un bacteri, un bacteri i un fong, una planta o un bacteri i un animal, diversos animals...; en una paraula, espècies de tots els regnes.
Altres relacions entre les espècies
Fins aquí hem considerat les relacions alimentàries (o tròfiques), però hi ha diferents menes d’interaccions interespecífiques (és a dir, entre espècies). En el paràgraf anterior s’ha parlat de la predació: es tracta d’una forma d’interacció interespecífica en què un animal es nodreix d’un altre animal. La competència, en canvi, es dóna quan dos organismes lluiten per alguna cosa que no és disponible en quantitat suficient per a tots dos. També s’observa en el món vegetal, on la competència per la llum, per exemple, empeny els arbres que creixen en grup a assolir alçades notables. Una altra interacció possible entre les plantes és l’antibiosi, és a dir, l’agressió d’una planta a una altra mitjançant excrecions químiques. Posem-ne dos exemples: el donzell obstaculitza el creixement del fonoll, i el rave impedeix que els créixens germinin.
El parasitisme és un tipus de relació en què un organisme (el paràsit) viu a la superfície o a l’interior d’un altre organisme, del qual obté les substàncies nutritives que necessita. Normalment, els paràsits estan molt "especialitzats" tant pel que fa a l’elecció de l’organisme hoste com a la seva estructura i cicle vital. El parasitisme és molt estès en la natura. Són paràsits tots els virus, molts bacteris, fongs i protozous, força plantes i un gran nombre d’animals. El parasitisme pot ser total o parcial: sense moure’ns del món vegetal, el vesc és un paràsit parcial de molts arbres i la cuscuta és un paràsit total del cànem i del llúpol.
La simbiosi és una relació interespecífica molt important en la qual els dos organismes que hi participen (els simbionts) obtenen avantatges (quan només un dels participants se’n beneficia es parla de comensalisme). Entre les plantes i els fongs sovint s’estableixen relacions d’utilitat recíproca: el bedoll, per exemple, proporciona substàncies orgàniques al fong Boletus scaber, que creix a les seves arrels, i el fong accelera l’absorció de nitrats per part del sòl. Els fongs tenen relacions simbiòtiques no tan sols amb les plantes, sinó també amb les algues blaves: els líquens són fongs en simbiosi amb algues blaves. D’altra banda, els fongs exerceixen l’antibiosi quant als bacteris: les floridures, formacions de fongs microscòpics, són productores actives d’antibiòtics. En parlar del cicle del nitrogen s’ha fet al·lusió a la simbiosi entre plantes i bacteris. S’estableix simbiosi entre les plantes que floreixen i els insectes pol·linitzadors: el benefici de l’insecte és l’aliment, i el de la planta, el fet de ser pol·linitzada.
Hi ha animals que viuen en simbiosi amb altres animals, de la mateixa manera que hi ha simbiosis entre animals i bacteris, entre animals i fongs, i entre animals i plantes. Hi ha formigues que viuen en simbiosi amb algunes espècies d’afídids: els deixen hibernar al formiguer, i a la primavera els transporten a les plantes de les quals els afídids són paràsits; en canvi, xuclen el líquid ensucrat secretat pels afídids. Molts mamífers allotgen bacteris a l’aparell digestiu. Els bacteris troben aliment en els aliments presents a l’estómac o a l’intestí i en canvi secreten productes útils per a l’hoste: es tracta d’una simbiosi entre animals i bacteris. En els humans és típic el cas dels bacteris que constitueixen la flora intestinal, indispensable per al procés de la digestió.
En general, es pot considerar el sistema viu com una immensa simbiosi entre espècies, encara que la relació entre individus no sigui simbiòtica. Un llop concret és predador d’una ovella concreta, però els llops com a espècie són d’alguna manera simbionts de l’espècie ovina ja que, en capturar més animals malalts que sans, exerceixen un control sobre els agents causants de malalties a les seves preses, a més de practicar una selecció de millora del seu patrimoni genètic.
La dinàmica de les successions ecològiques
No tan sols s’estableixen relacions entre espècies presents simultàniament en una certa regió (coetànies), sinó també entre espècies que apareixen en aquesta mateixa regió però en diferents temps, segons una successió no casual. La successió de les espècies constitueix el desenvolupament de l’ecosistema local, que segueix un ritme global caracteritzat, al començament, per un clar predomini de la fotosíntesi sobre la respiració i, per tant, per una acumulació de biomassa. Durant aquesta fase es produeix una ràpida substitució de les espècies. Posteriorment es crea una associació d’espècies que es caracteritza per l’estabilitat, i aleshores es diu que l’ecosistema ha assolit el clímax. En aquesta fase, el volum total de la fotosíntesi disminueix (com també disminueix el volum total de la respiració), de manera que fotosíntesi i respiració s’equilibren. El desenvolupament de l’ecosistema planetari també ha seguit —durant milers de milions d’anys— un ritme en el qual ha predominat la fotosíntesi sobre la respiració, i després ha deixat pas a un equilibri substancial (vegeu "Breu història del sistema viu"). Però, com és que al cap de milers de milions d’anys del començament de la vida al nostre planeta hi ha ecosistemes que encara no han assolit el clímax? Per explicar aquest fet, i per sintetitzar el procés de les successions ecològiques que poden portar un ecosistema a una situació de clímax, partim d’una catàstrofe natural, com una erupció volcànica o un incendi. Encara que la catàstrofe deixi la roca al descobert, la vida pot recomençar. Com hem vist a "La vida, el cicle de l’aigua i la llum", les plantes creixeran en qualsevol escletxa de la roca on hagi quedat una mica d’aigua, i amb les seves excrecions àcides eixamplaran les escletxes, les quals encara s’eixamplaran més quan l’aigua que contenen es glaci. Amb la fragmentació de la roca s’acompleix el primer pas cap a la formació de la base mineral de l’humus. S’hi faran llavors invisibles, espores bacterianes, i els organismes que en naixeran deixaran detritus i despulles que començaran a formar la substància orgànica de l’humus, i les associacions de vegetals se succeiran, cada vegada més exigents quant a la presència d’humus, però diferents d’un lloc a un altre segons les condicions climàtiques i edàfiques.
En tot cas, però, es tractarà d’una repoblació per part d’organismes expulsats del seu lloc d’origen: espores portades pel vent, llavors transportades pels insectes o els ocells, però no es formarà vida de la no-vida, com va passar fa 3 500 milions d’anys, o potser més. El naixement de la vida a partir de la no-vida només s’ha esdevingut una sola vegada, i no es pot repetir perquè la vida mateixa ha modificat el planeta. L’ha fet més acollidor per a la vida, però absolutament inhòspit per al naixement de la vida a partir de la matèria inerta.