Dels animals unicel·lulars als pluricel·lulars
Una característica essencial dels éssers vius o organismes és l’intercanvi de substàncies amb el món exterior. Per un animal o una planta, viure significa apropiar-se d’alguna cosa que no és al seu interior. Aquesta cosa pot ser l’oxigen, el diòxid de carboni de l’aire o de l’aigua, les sals minerals del sòl o de l’aigua, o substàncies complexes com els sucres, els greixos, les proteïnes i les vitamines. Les plantes verdes fabriquen, per l’acció de la llum solar, les substàncies que els són necessàries per al metabolisme. Són laboratoris complexos on contínuament es realitzen combinacions químiques. Els animals, en canvi, obtenen les substàncies nutritives menjant-se les plantes (els herbívors) o menjant-se els herbívors (els carnívors), i també n’hi ha que són paràsits de plantes o d’altres animals.
La diferència fonamental entre els organismes vegetals i animals és, doncs, el fet que els primers es nodreixen a partir de matèria inorgànica, mentre que els segons necessiten l’aportació de substàncies produïdes per altres organismes. És per això que els animals reben el nom d’heteròtrofs (a partir de dos mots grecs que signifiquen "nodrir-se d’altres") i viuen gràcies a l’existència d’organismes autòtrofs ("que es nodreixen per ells mateixos").
Un cop esbossada la principal diferència entre el món animal i el vegetal, ens disposem a descriure l’ampli i variat conjunt dels organismes animals i ho fem a partir d’una classificació dels diversos grups, seguint un ordre creixent de complexitat de l’estructura corporal i de les funcions vitals.
Els protozous són eficaços
El medi que ens envolta és ple d’organismes eucariotes heteròtrofs que ens passen desapercebuts, ja que la seva petitesa (llevat d’algunes excepcions) els fa invisibles a ull nu. Es tracta dels protozous, organismes unicel·lulars. En les classificacions modernes, es considera que els protozous pertanyen a un regne específic, el dels protoctists. Tradicionalment, però, s’incloïen dins el regne dels animals perquè hi tenen moltes característiques comunes. D’altra banda, la validesa del regne dels protoctists és encara qüestionada per alguns estudiosos, que prefereixen mantenir-se fidels a les classificacions que no preveuen aquest regne. Seguint la posició clàssica —possiblement en vies d’extinció— comencem el capítol relatiu al regne animal precisament amb aquests organismes microscòpics, i la raó són les afinitats que els relacionen amb els "veritables" animals.
En els animals unicel·lulars totes les funcions característiques de l’organisme viu, com la nutrició, el creixement i la reproducció, són dutes a terme per una sola cèl·lula totalment autosuficient. En els animals pluricel·lulars, en canvi, són a càrrec de l’acció coordinada i interdependent de milers de milions de cèl·lules.
Si examinem un organisme unicel·lular, encara que sigui particularment evolucionat com el parameci, resulten sorprenents la varietat i l’eficàcia de les seves estructures i les adaptacions que aquesta única cèl·lula realitza per a aconseguir satisfer totes les necessitats vitals.
La cèl·lula és revestida per una pel·lícula elàstica, però consistent, que li permet conservar la forma. A la superfície externa hi ha cilis que configuren l’estructura de locomoció. La membrana cel·lular presenta dues obertures que fan de boca i anus, i la digestió és segregada a l’interior d’un orgànul especial, el vacúol digestiu. A més, pot mantenir constant la composició interna per mitjà dels vacúols (és a dir, petites cavitats delimitades per la membrana) contràctils que expulsen l’aigua sobrera. Fins i tot disposa d’un esbós, per bé que molt primitiu, del sistema nerviós.
Tot i que es tracta d’un organisme autosuficient i organitzat, presenta alguns desavantatges respecte dels organismes formats de moltes cèl·lules. En primer lloc, és molt més vulnerable. Qualsevol part danyada afecta tot l’organisme i per tant és molt més difícil de guarir; i, sobretot, les seves possibilitats de creixement són molt limitades.
Des del punt de vista evolutiu, la necessitat de tenir una massa corporal més voluminosa va afavorir el desenvolupament de formes pluricel·lulars, a partir de l’organisme progenitor format per una sola cèl·lula. Efectivament, els organismes més grossos poden fer moltes més coses que els éssers més petits. A més, l’augment de les dimensions corporals ofereix noves alternatives: nous mecanismes d’alimentació, nous sistemes de locomoció i nous hàbitats per explotar. Molts organismes pluricel·lulars són fins i tot capaços de regenerar parts del seu cos en el cas de ser mutilats (vegeu "Regeneració i autotomia").
Al costat d’aquestes oportunitats, però, també apareixen problemes: l’augment de les dimensions implica la presència d’una estructura de sosteniment i, sobretot, d’un sistema de difusió i transport de les diverses substàncies per l’interior del cos.
Regeneració i autonomia
Una de les facultats més meravelloses que tenen els éssers vius és la possibilitat de reconstruir íntegrament els teixits i òrgans que han patit lesions. És un fenomen estès en els animals i els vegetals, que inclou processos de cicatrització, sempre que les lesions no siguin gaire greus. Però en alguns organismes aquesta capacitat es manifesta en grau màxim en obrar veritables miracles en la reconstrucció ex novo de parts senceres del cos, i també de tot l’organisme a partir d’un fragment petit. Són els casos de regeneració que presenten les plantes i alguns animals. Pel que fa als animals, aquest fenomen és més freqüent entre els invertebrats, tot i que també hi ha exemples coneguts entre els vertebrats. Un trosset del cos d’una planària (cuc pla que pobla les aigües dels nostres rius i llacs), sempre que no sigui massa petit i es conservi en aigua en bones condicions, ben aviat tornarà a adquirir la forma d’un individu sencer, reformant la seva substància, reconstituint els òrgans que li falten i creant nous teixits a partir del punt on s’ha produït el tall.
Un braç d’una estrella de mar arrencat de la resta del cos regenera el disc central i els altres quatre braços. Aleshores apareix una curiosa forma de cometa, que després es redueix i es regularitza gradualment, per bé que es manté força temps la desproporció entre el braç originari i les parts regenerades. Certs animals d’organització més complexa també poden presentar aquests fenòmens: nombrosos crustacis, alguns insectes i diverses espècies d’aranyes regeneren els apèndixs (les antenes i les parts de l’aparell bucal) perduts o tallats.
De vegades, però, la part que es regenera a conseqüència d’amputacions és molt diferent de la perduda. En les gambes, per exemple, es pot formar una antena al lloc d’un ull pedunculat. Entre els amfibis, els capgrossos de les granotes i els gripaus, dels tritons i de les salamandres, i fins i tot els adults, són capaços de regenerar la cua o les potes. Entre els rèptils, és ben conegut que les sargantanes poden regenerar la cua. No és gens estrany trobar una sargantana amb la cua parcialment regenerada, o amb l’extrem terminal doble, a causa d’un procés regeneratiu anormal.
En diversos animals pertanyents als grups zoològics més diversos, entre els quals les sargantanes, els crancs i les aranyes, el poder de regeneració es relaciona amb la facultat de desprendre’s espontàniament d’algunes parts del cos (autotomia) amb finalitat defensiva. Així, una sargantana que és aferrada per la cua per un predador, perd la cua i s’escapa. Si s’atrapa una gamba, un cranc o una aranya per una pota, sovint el membre es trenca i l’animalet fuig.
En aquests casos es tracta d’un reflex provocat per la captura (però també per un fort augment de la temperatura), que es manifesta amb una contracció instantània molt enèrgica dels músculs situats al punt on es verifica la ruptura.
Hi ha petits rosegadors (els lirons i alguns ratolins) que tenen una reacció semblant: retinguts i agafats per l’extrem de la cua, abandonen l’embolcall cutani. El tros de la cua, ja despullat, cau tot seguit i l’animal regenera l’extremitat amb el plomall de pèls característic.
La colònia de cèl·lules
Una manera de solucionar la manca d’augment global de les dimensions de l’organisme és la colònia cel·lular. S’origina a partir de la unió de moltes cèl·lules que, tot i estar estretament interconnectades, mantenen la seva individualitat. Si una colònia és fragmentada, les diverses parts poden sobreviure i ser independents. Al principi les colònies eren simplement un aglomerat d’organismes unicel·lulars units per lligams més aviat làbils. Després, probablement per una forta pressió selectiva dirigida a millorar l’eficàcia de tot el sistema, es va començar a crear una mena de "divisió del treball" que cada vegada era més pronunciada. Totes les cèl·lules de la colònia, a mesura que assumien una tasca concreta, entraven en un procés de diferenciació i especialització i deixaven de ser un individu casualment associat a altres per a esdevenir part integrant d’un nou organisme més complex. Si es fes un estudi de totes les colònies cel·lulars que hi ha actualment, s’observarien infinites gradacions entre les dues situacions extremes: des de la simple agregació d’organismes unicel·lulars, cadascun amb la seva pròpia individualitat i units d’una manera aparentment casual, fins a colònies molt properes a les condicions de pluricel·lularitat, on grups de cèl·lules ben diferenciades fan les diverses funcions d’una manera "especialitzada".
Un exemple de colònia d’aquest darrer tipus són les esponges, entre les quals hi ha grups de cèl·lules encarregades de funcions específiques, reunides en teixits. Algunes constitueixen un teixit extern de protecció, d’altres entapissen la cavitat interna i absorbeixen els aliments que després passaran a les cèl·lules dels altres teixits i n’hi ha que fabriquen petits elements esquelètics per sostenir tota l’estructura. La pluricel·lularitat de les esponges és encara molt rudimentària; les cèl·lules s’associen d’una manera poc rígida i es poden desprendre fàcilment les unes de les altres i formar altres aglomerats independents.
Els organismes pluricel·lulars
Les cèl·lules dels organismes pluricel·lulars (amb l’excepció dels òvuls de molts animals, que tenen certa autonomia) no poden sobreviure aïllades perquè formen part d’un sistema basat rígidament en la llei de la divisió de les funcions. Les primeres cèl·lules d’un organisme, les cèl·lules embrionàries, no presenten diferències entre elles, almenys exteriorment, però en esdevenir adultes pateixen un procés profund de diferenciació. Cada tipus cel·lular es modifica i s’especialitza per a la funció concreta que ha de desenvolupar. Per exemple, les cèl·lules musculars esdevenen molt allargades i capaces de contreure’s, les cèl·lules nervioses desenvolupen unes prolongacions primes i s’especialitzen en la conducció d’estímuls, i les cèl·lules de revestiment esdevenen aplanades i s’uneixen estretament de manera que constitueixen una eficaç capa protectora. L’autonomia dels organismes unicel·lulars es perd i cada cèl·lula ha de subordinar les pròpies activitats vitals a la tasca que se li encarrega, però a canvi pot confiar en l’ajut complementari de les altres cèl·lules especialitzades. El resultat global és un gran augment de l’eficàcia de tot el sistema.
Els principals models de l'estructura corporal dels animals
Els animals, si bé presenten diferències que en alguns casos semblen abismals, són en realitat menys diferents del que aparenten. Dins dels grups zoològics principals, la suma dels quals és un nombre limitat, hi ha una unitat substancial d’estructura que també és detectable sota el que podrien semblar les variacions més fantàstiques. Això és força evident si es té en compte que totes les espècies que pertanyen a cadascun dels grans grups (phyla, singular phylum; a partir d’ara parlarem de divisió) en què es classifiquen els organismes provenen d’avantpassats comuns (pel que fa als problemes de la classificació, vegeu "Vida i evolució"). Ara ens ocuparem de les principals divisions confirmades amb més èxit al llarg de l’evolució dels éssers vius. Algunes d’aquestes divisions són tan semblants, que en les classificacions anteriors apareixien unificades.
Els porífers
Els porífers, o esponges, es consideren els animals pluricel·lulars vivents més arcaics, tot i que, com s’ha dit abans, l’organització de les seves cèl·lules en teixits és encara molt rudimentària.
Les esponges tenen formes i dimensions extremament diverses (i sovint colors molt bonics), però una estructura fonamental més aviat simple: un sac recorregut per molts canalicles que comuniquen amb l’exterior per mitjà de porus (d’on ve el nom del grup). A través d’aquestes obertures, l’aigua marina on viuen els porífers pot penetrar a l’interior de l’organisme fins a la cavitat interna i assegurar així els intercanvis gasosos i el subministrament de substàncies alimentàries.
A la paret de la cavitat interna hi ha unes cèl·lules específiques, especialitzades en la "captura" i absorció de l’aliment, mentre que una altra mena de cèl·lules formen el teixit de revestiment extern.
A més, les esponges tenen un esquelet rígid format per molts elements cornis minúsculs (o calcaris o silícics). Les "esponges" naturals emprades normalment en la higiene humana són precisament els esquelets cornis dels porífers.
Els celenterats
Els celenterats són els primers organismes amb una organització pluricel·lular pròpiament dita. En efecte, apareixen amb diversos teixits diferenciats. Encara que el terme celenterat tradicionalment ha reunit els cnidaris i els ctenòfors, dos grups que actualment es considera que han aparegut i evolucionat separadament, aquí parlarem de celenterats per referir-nos tan sols als cnidaris, segons la classificació clàssica.
Pertanyen a aquest grup les meduses, els coralls, les hidres i les anemones de mar. Són organismes amb una estructura simple però particularment aconseguida, tal com ho prova la seva difusió, ja que els celenterats són un dels grups animals més nombrosos. Es tracta essencialment d’un sac revestit a l’interior per una capa contínua de cèl·lules, l’endoderma, i a l’exterior per un altre teixit amb funció protectora.
Els intercanvis amb el medi tenen lloc per mitjà d’una sola obertura, a través de la qual s’introdueixen els aliments i s’expulsen els residus. La boca i l’anus no es diferencien.
Mentre que alguns celenterats com les actínies i els pòlips colonials (com els coralls i les madrèpores) es mantenen enganxats al fons marí durant la major part de la seva vida, d’altres, com les meduses, són nedadors actius. En aquest últim cas el cos adopta forma d’ombrel·la, amb una membrana que circumda la boca. La medusa es mou aprofitant les contraccions que recorren aquesta membrana.
Els celenterats són els únics animals pluricel·lulars de simetria radiada tant externa com interna; és a dir, que el cos pot ser dividit en dues parts iguals per un nombre elevat de "plans" disposats radialment respecte d’un eix. En general, els altres animals presenten en alguns casos una simetria bilateral (hi ha un sol pla que divideix el cos en dues meitats iguals) i en altres casos l’estructura externa permet traçar diversos plans de simetria, però els òrgans interns no respecten aquesta simetria; és el que passa en les garotes i les estrelles de mar. Aquesta distinció és molt important amb vista a la classificació.
Pòlips o meduses?
Els pòlips són organismes marins que pertanyen als celenterats. En les colònies més evolucionades de pòlips, com que no és indispensable que cada individu busqui l’aliment pel seu compte, alguns d’ells s’especialitzen en altres funcions. Uns pòlips es dediquen a la reproducció i d’altres adopten una simple forma allargada, es recobreixen d’una capa espessa de cèl·lules urticants i es dediquen només a funcions defensives. La reproducció es duu a terme amb la formació de gemmes sexuals proveïdes de molts braços, que recorden un pop petit. Però en comptes de créixer seguint l’estructura habitual, la gemma s’expandeix en forma d’ombrel·la i en desprendre’s comença a nedar com si fos un pop de cap per avall, de forma més aviat aplanada: és la medusa. La medusa no és sinó una de les dues formes biològiques, una de les dues maneres de ser dels celenterats. Quan es forma, no és gaire diferent d’un pòlip, però a poc a poc esdevé ampla, lleugera i adaptada a la natació.
Cal explicar per què de vegades els celenterats presenten l’estadi de medusa. La manera més comuna i immediata que té un pòlip de reproduir-se és l’anomenada "gemmació"; al peu d’un individu apareix una gemma que ben aviat esdevé un nou pòlip. Aquest tipus de reproducció és molt ràpida, simple i directa, i garanteix un nombre notable de nous individus. Però té un inconvenient: els nounats són iguals que el progenitor, del qual no difereixen des del punt de vista genètic. La reproducció sexual elimina aquest inconvenient i permet la formació de les futures generacions recombinant els caràcters hereditaris dels pares.
Així, si el sexe es consolida en resposta a l’exigència de produir la varietat més àmplia possible de caràcters, és evident que això requereix la barreja (= recombinació) de patrimonis genètics de poblacions que estiguin allunyades. Per tant, cal que almenys un estadi de la vida de l’individu sigui mòbil, per tal que es pugui desplaçar. L’estadi mòbil dels celenterats correspon precisament a la larva i la medusa.
Hi ha diverses solucions, dins l’àmbit dels celenterats, quant a la forma predeterminada d’existència. Alguns, els hidrozous, sovint presenten un cicle regulador pòlip/medusa: en aquest cas es tracta de meduses petites (el diàmetre sol ser d’1 cm) i de pòlips (d’1-2 mm a pocs centímetres) que normalment s’agrupen en colònies. En canvi, hi ha altres celenterats com els escifozous en què predomina l’estat de medusa: el pòlip és molt petit i aviat se segmenta per a generar petites meduses que després creixen i formen meduses de grans dimensions. La medusa més gran que es coneix, la Cynea árctica, té una ombrel·la de 2 m de diàmetre i tentacles de fins a 40 m de llarg. L’última classe dels celenterats, la dels antozous, es caracteritza, en canvi, per l’exclusivitat de la forma polipoide: els pòlips solitaris (com les actínies o anemones de mar) o els pòlips colonials (com els coralls i les madrèpores).
Els cucs plans (platihelmints)
Són bàsicament dos els elements que distingeixen els cucs plans dels altres grups de cucs: la forma aplanada, que els fa semblants a una cinta, i l’aparell digestiu. Com en els celenterats, l’aparell digestiu consta d’una sola obertura situada al costat ventral. A més, la cavitat digestiva, que té el fons cec, s’estén ramificant-se per quasi tot el cos de l’animal, de manera que queda assegurada una distribució directa de les substàncies nutritives. En efecte, els cucs plans no tenen líquid circulant ni aparell circulatori. En tots els altres grups animals, en canvi, hi ha dues obertures diferenciades, la boca i l’anus, i el sistema digestiu es configura com un tub més o menys especialitzat.
Encara que s’observin amb atenció, no és fàcil reconèixer el cap d’aquests animals: s’aprecia més fàcilment quan l’animal es desplaça, perquè és la part del cos que es mou primer. Anatòmicament, però, és ben caracteritzat perquè presenta una certa concentració de teixit nerviós. Els òrgans sensorials, per exemple les cèl·lules fotosensibles, normalment també s’agrupen a l’extremitat anterior.
Molts platihelmints són paràsits i viuen a l’interior d’altres animals. Per aquest motiu presenten regressions i modificacions d’òrgans típiques que els distingeixen dels platihelmints "lliures". Els paràsits més coneguts són les tènies, de fins a 6 m de llarg, que s’instal·len a l’aparell digestiu dels vertebrats, on s’enganxen a la paret intestinal de l’hoste. Manquen completament d’aparell digestiu i, com que viuen en un medi saturat d’aliment gairebé del tot digerit, poden obtenir tot el nodriment que necessiten absorbint-lo directament a través de la superfície cutània. A més, com a adaptació a la vida parasitària no posseeixen òrgans fotosensibles, però tenen la funció reproductiva molt desenvolupada: el cos és dividit en segments i els que estan més allunyats del cap esdevenen veritables sacs plens d’ous madurs que es desprenen i són emesos a l’exterior.
Els cucs cilíndrics (nematohelmints)
El grup de nematohelmints és constituït per animals sobretot aquàtics. Els terrestres menen vida activa solament al sòl o en zones humides; molts són paràsits de l’espècie humana, els animals i les plantes. En la fase de latència, quan l’organisme suspèn l’activitat vital per les condicions ambientals adverses, poden suportar períodes llarguíssims de dessecació quasi completa.
La divisió més significativa és la dels nematodes. Tenen el cos cilíndric, amb un cap ben diferenciat, i l’aparell digestiu és representat per un tub que parteix de la boca i acaba a l’anus.
Molts cucs cilíndrics presenten, entre la paret externa i el tub digestiu, una cavitat més aviat gran i plena de líquid, i al seu interior les diverses substàncies circulen d’un extrem a l’altre del cos. Aquesta cavitat no comunica ni amb l’exterior ni amb l’aparell digestiu i conté suspesos els òrgans reproductors i el tub digestiu, que d’aquesta manera tenen certa llibertat de moviment quan l’animal es plega, s’arrossega o neda.
Per aquesta característica, els cucs cilíndrics constitueixen un estadi intermedi entre els platihelmints, que per paret tenen una estructura compacta i sense llacunes, i els anèl·lids, que presenten una veritable cavitat en la qual els òrgans viscerals s’allotgen fermament.
Els anèl·lids
Pertanyen a aquesta divisió tant els cucs terrestres (el cuc de terra típic) com els aquàtics (d’aigua dolça i marina). Alguns, a més, són paràsits. Els anèl·lids posseeixen una característica important, la metameria: el cos, segmentat, es compon d’una sèrie d’unitats cilíndriques iguals (tret del cap), amb òrgans i estructures biològiques repetides diverses vegades. Alguns animals més evolucionats presenten vestigis d’organització metamèrica; només cal pensar en les vèrtebres i els nervis espinals dels vertebrats.
En els anèl·lids, cada segment conté una cavitat plena de líquid, el celoma, on hi ha suspesos per mitjà de membranes el sistema digestiu (un tub que recorre tot el cos de l’animal, de la boca a l’anus, amb zones especialitzades en les diverses funcions digestives) i els altres òrgans. Apareix per primera vegada un veritable sistema circulatori.
El sistema nerviós també presenta cert grau d’especialització i el cos és enterament recorregut per una cadena ganglionar amb estructura metamèrica; un parell de ganglis per segment.
Alguns anèl·lids tenen apèndixs que tant serveixen per a moure’s a la terra ferma com per a nedar.
Els artròpodes
Els artròpodes són una de les divisions que han tingut més èxit en el poblament de la Terra. S’ha calculat que aproximadament el 80% de totes les espècies existents pertanyen a aquest grup. Evidentment, un model d’organització com aquest, amb un cos dividit en segments i òrgans especialitzats per a les diverses funcions, s’ha revelat particularment reeixit, versàtil i eficaç. Presenta moltes analogies amb l’estructura dels anèl·lids, però els artròpodes tenen a més un esquelet extern (l’exosquelet), rígid i flexible alhora, que fa de sosteniment i protecció. Les extremitats també hi són presents, en general més aviat complexes. Algunes han perdut la funció locomotora i serveixen per a la nutrició i la reproducció. La cavitat del cos dels artròpodes és molt reduïda i conté l’hemolimfa, un líquid amb les mateixes funcions que la sang dels vertebrats, que circula per l’acció del cor. El sistema nerviós, particularment complex i perfeccionat, és constituït per fibres i ganglis.
Després d’il·lustrar breument les característiques generals d’aquesta divisió ens entretindrem una mica més en els seus grups i classes principals:
a) Els crustacis. El cos dels crustacis, compost per segments diferents, és constituït per 3 regions: el cap, el tòrax i l’abdomen. El cap, sovint unit al tòrax, amb el qual forma el cefalotòrax, té un parell de mandíbules, 2 parells de maxil·les, 2 parells d’antenes i un parell d’ulls compostos o simples, pedunculats o no. Els apèndixs del tòrax constitueixen les potes, i alguns dels parells més anteriors funcionen com a òrgans prensors i mastegadors de l’aliment. Normalment el primer parell de potes acaba en unes pinces robustes. Els segments de l’abdomen són mòbils i tenen apèndixs reduïts, de vegades absents. L’exosquelet, impregnat de calci, és particularment dur i resistent.
Els crustacis es distingeixen dels altres artròpodes essencialment per dos elements: tenen 2 parells d’antenes i respiren exclusivament per mitja de brànquies. Viuen a l’aigua, tant dolça com salada, i poden tenir dimensions molt diferents, que van del mil·límetre de la puça d’aigua als 50 cm de la llagosta.
b) Els aràcnids. La classe dels aràcnids agrupa animals molt coneguts com els escorpins, les aranyes, les paparres (àcars) i altres grups. Són animals generalment terrestres, caracteritzats per tenir un sol parell d’apèndixs davant l’obertura bucal, a vegades dotats de pinces en forma de tenalla per agafar i trossejar les preses. La segmentació del cos és poc evident. El cap i el tòrax estan fusionats en el cefalotòrax, i l’abdomen forma sovint un sac sense rastre de segmentació. En el cefalotòrax hi ha 6 parells d’apèndixs, els 2 primers modificats per a realitzar les funcions nutritiva i tàctil. Molts tenen glàndules verinoses (els escorpins i les aranyes) i altres posseeixen glàndules sericígenes especials, mitjançant les quals teixeixen les teranyines i fabriquen els capolls.
c) Els miriàpodes. El grup dels miriàpodes (que reuneix 4 classes) és constituït per petits animals amb el cos subdividit en molts segments iguals, cadascun dels quals dotat d’un parell de potes. L’exemple més representatiu d’aquesta classe és el centpeus.
d) Els insectes. Són els animals més difosos del món. Se n’han classificat 800 000 espècies, però sembla que com a mínim encara n’hi ha 1 milió i mig més de desconegudes. Actualment a la Terra hi ha aproximadament un milió de bilions d’insectes i s’ha calculat que la proporció entre l’espècie humana i aquests animals és d’uns 200-300 milions d’insectes per ésser humà. L’element que ha determinat aquesta enorme difusió ha estat probablement l’adaptació al vol, que ha permès l’explotació de tots els hàbitats del planeta.
Els insectes tenen un exosquelet robust però lleuger i flexible, 3 parells de potes i un cos subdividit en 3 regions: el cap, el tòrax i l’abdomen. El cap incorpora un parell d’antenes i un aparell bucal complex que, segons el tipus d’alimentació, és diversament modificat. L’ordre dels himenòpters (les abelles i les formigues) i el dels lepidòpters (les papallones), per exemple, tenen un aparell bucal xuclador. L’ordre dels coleòpters (els borinots, les marietes i els escarabats) i el dels ortòpters (les llagostes) tenen un aparell mastegador. L’ordre dels hemípters (les xinxes i les cigales), el dels sifonàpters (les puces) i el d’alguns dípters (les mosques i els mosquits) són dotats, en canvi, d’un sistema que és alhora punxant i xuclador.
El tòrax és format per 3 segments que gairebé sempre estan fixats i soldats, i cadascun inclou un parell de potes. El segon i el tercer segments incorporen, a més, un parell d’ales cadascun (alguns paràsits com les puces les han perdudes), constituïdes per dues membranes quitinoses. En alguns casos només un parell d’aquestes ales és funcional. En els coleòpters, per exemple, les ales anteriors formen una mena d’escut rígid que protegeix les ales posteriors, mentre que en els dípters, com els mosquits, les mosques, etc., són les posteriors que es transformen en òrgans d’equilibri.
L’abdomen no té apèndixs, és molt mòbil i comprèn com a màxim 11 segments units per membranes toves.
Els insectes respiren per mitjà de tràquees i tenen un sistema nerviós amb ganglis grossos que ja és força evolucionat.
Entre els insectes és molt estès el fenomen de la metamorfosi. Els individus acabats de néixer arriben a la forma adulta a través de diverses transformacions que comporten canvis profunds de l’estructura anatòmica i dels hàbits de l’animal (vegeu "La metamorfosi").
Els mol·luscs
La divisió dels mol·luscs comprèn animals extremament diferents i amb diversos graus de complexitat. Només cal pensar en les diferències que s’observen entre un cargol, una sípia i una ostra. Però tots tres, i la resta de mol·luscs, tenen en comú un mateix model estructural: un cos compacte, on els apèndixs no estan aparellats ni sobresurten dels segments corporals, dotat de peu muscular, mantell i conquilla. El peu es caracteritza per una sòlida massa muscular, serveix per a la locomoció i pot ser molt modificat, fins al cas extrem dels cefalòpodes (els pops, les sípies i els calamars), en què s’ha transformat en un conjunt de tentacles al voltant de la boca.
El mantell és com un sac que conté les vísceres i s’encarrega de la secreció de la conquilla, que pot ser gran i elaborada, cas en què té funció protectora, tal com s’observa en els mol·luscs bivalves (els musclos i les ostres) i els gastròpodes (els cargols, els llimacs...), o bé pot ser petita i interna i funcionar essencialment com a òrgan de sosteniment, com és el cas de la majoria dels cefalòpodes (les sípies i els calamars), i fins i tot pot ser del tot absent, com passa en els pops i els gastròpodes nudibranquis.
Tots els mol·luscs, excepte els gastròpodes terrestres, respiren per mitjà de brànquies. Tenen un sistema nerviós força desenvolupat, i especialment perfeccionat en els cefalòpodes. Tenen també els ulls molt evolucionats. El cervell dels pops, quant a la complexitat estructural i funcional, és fins i tot comparable al dels vertebrats.
Els equinoderms
Els representants més coneguts d’aquesta divisió són les estrelles de mar, les garotes i les holotúries (anomenades "cogombres de mar"). Comprèn organismes molt diversos que tenen en comú una sola característica: la simetria pentaradial. És a dir, el cos d’aquests animals es pot dividir en 5 parts iguals per mitjà de 5 plans.
Alguns òrgans interns també presenten simetria d’aquesta mena. El sistema nerviós, per exemple, és constituït per un anell que circumda la boca i del qual surten 5 nervis radials. En les estrelles de mar, l’estómac es divideix en 5 branques que es prolonguen a l’interior de cada braç. Igualment mereix ser esmentat el sistema locomotor dels equinoderms, ja que es mouen per mitjà de petits pedicels plens de líquid que s’allarguen i es contreuen alternativament.
Sorprenentment, aquest grup és el més proper als cordats, divisió a la qual pertanyen els vertebrats. Entre els animals adults de tots dos grups no hi ha cap mena de semblança, però les formes embrionàries tenen molts trets comuns que suggereixen la hipòtesi d’un avantpassat comú relativament proper.
Els vertebrats
El model d’organització corporal dels vertebrats és un dels que han tingut més èxit des del punt de vista evolutiu. Constitueixen un grup nombrós que, com els artròpodes, no tan sols ha conquerit tots els ambients terrestres i aquàtics, sinó també l’aire.
La característica més evident d’aquest model estructural és la presència d’un esquelet intern (l’endosquelet). En la majoria dels vertebrats és format per un material únic en el seu gènere, el teixit ossi, produït per la secreció de milers de cèl·lules especialitzades. Els ossos que en resulten són particularment rígids i durs, però estan articulats de tal manera que proporcionen una estructura extremament flexible que permet gran varietat de moviments. Per a la locomoció, de l’eix de sosteniment principal surten 4 extremitats, que al seu torn disposen d’esquelet intern articulat. Malgrat que en alguns grups s’han modificat profundament (en les serps fins i tot han desaparegut), en tots els grups es pot reconèixer la mateixa estructura fonamental. La part dorsal de l’esquelet, la columna vertebral, és segmentada i recorreguda per un canal que conté la medul·la espinal. Aquest cordó nerviós és situat en posició dorsal respecte al canal digestiu, i no pas ventral com en els altres grups.
En general, el sistema nerviós dels vertebrats és el més complex i especialitzat de tots els grups animals. Té particularment desenvolupada la regió destinada a la integració i la coordinació dels impulsos nerviosos, és a dir, el cervell. Els òrgans sensorials també són molt perfeccionats: els vertebrats tenen en general capacitats olfactives, auditives, visuals... molt superiors a les dels invertebrats.
Una altra característica d’aquest grup és la complexitat del sistema circulatori tancat (és a dir, que la sang sempre roman a l’interior dels vasos), format per un cor i una sèrie de vasos.
Els vertebrats són en realitat una subdivisió dels cordats, juntament amb dos altres grups animals molt simples: els cefalocordats i els urocordats (un cefalocordat típic és l’amfiox, una mena de peixet minúscul). Els cordats, encara que són animals molt diferents, comparteixen la presència, durant la vida embrionària, d’un cordó nerviós dorsal, de fissures branquials i d’un òrgan de sosteniment especial, el cordó dorsal. Els cefalocordats conserven aquestes estructures tota la vida. Els urocordats (grup al qual pertanyen els ascidis, que superficialment s’assemblen d’una manera notable a les esponges), quan són adults, tan sols conserven els forats branquials. Entre els vertebrats es conserva el cordó nerviós dorsal un cop superat l’estat embrionari, mentre que solament els adults d’algunes classes (els peixos i alguns amfibis) conserven els forats branquials.
El grup dels vertebrats comprèn diferents classes: els peixos, els amfibis, els rèptils, els ocells i els mamífers.
a) Els mamífers, excepte els més primitius (els monotremes, que ponen ous, i els marsupials, que pareixen embrions encara incomplets; vegeu més endavant "L'ou"), pareixen cries vives i ja formades, a les quals alleten i dediquen especial atenció, que a vegades es manté força temps. Són homeoterms, és a dir mantenen una temperatura interna constant, gràcies a la contribució d’un revestiment protector de pèls. A més, tenen una dentadura òssia molt desenvolupada i diferenciada que depèn del tipus d’alimentació, i un cervell perfeccionat, en el qual són particularment refinats els processos mentals d’integració i coordinació.
b) Els ocells posseeixen una estructura altament especialitzada per al vol. Les extremitats anteriors s’han modificat i han originat les ales, l’estern ha adoptat la forma d’una làmina òssia i la majoria dels ossos han esdevingut buits i molt lleugers. El sistema respiratori s’ha modificat i perfeccionat fins a assolir una eficàcia extrema. Com els mamífers, els ocells també són homeoterms i tenen el cos recobert d’una capa protectora i aïllant, en aquest cas constituïda per plomes. La boca és un bec i no té dents.
c) Els rèptils, en canvi, són poiquiloterms, és a dir que la seva temperatura interna no és constant, sinó que varia en funció de les condicions ambientals. Tenen el cos recobert d’escames o plaques còrnies, les quals es renoven periòdicament amb el procés de la muda. La boca presenta unes dents no gaire especialitzades. Com els ocells, ponen ous protegits per una closca dura, on els embrions es poden desenvolupar lluny dels perills ambientals i, sobretot, del risc de l’evaporació. Els rèptils comprenen diversos ordres, entre els quals hi ha: els quelonis (les tortugues), amb el cos protegit per la closca òssia; els crocodilians (cocodrils), que tenen un cos de grans dimensions revestit de plaques òssies i cua aplanada, i els escatosos, que inclouen els saures (les sargantanes i llangardaixos, els camaleons i les iguanes), quasi sempre dotats de potes, i els ofidis (les serps), sense extremitats i amb el cos molt allargat.
d) Els amfibis viuen tant a terra ferma com a l’aigua i són un grup de transició entre els animals exclusivament aquàtics i els terrestres. El seu cicle vital se subdivideix en dues fases: un primer període en què s’assemblen als peixos i una fase adulta en què les característiques són més orientades cap a una adaptació al medi terrestre. Les espècies que viuen predominantment a terra ferma, però, també mantenen una relació de dependència amb l’aigua: quan es reprodueixen tornen al medi aquàtic per posar-hi els ous i fecundar-los. Entre els amfibis es distingeixen: els urodels, de cos allargat, amb la cua llarga i les potes curtes (les salamandres, els tritons i els axolots), i els anurs, de cos massís i sense cua, però amb les potes posteriors llargues i robustes, particularment apropiades per a saltar i nedar (les granotes i els gripaus).
e) Hi ha dues classes de peixos : els condrictis o peixos cartilaginosos com els taurons i les rajades, etc., és a dir, els anomenats peixos bestinals o bestines, i els osteïctis o peixos ossis, que són els més difosos (els peixos pròpiament dits). Són organismes completament adaptats a la vida aquàtica, no tenen potes sinó dos parells d’aletes parelles (pectorals i ventrals), a més d’aletes imparelles situades a l’extrem dorsal i ventral del cos i a la cua. No posseeixen pulmons, sinó brànquies a través de les quals capten l’oxigen dissolt en l’aigua.
Antigament també es consideraven peixos les espècies pertanyents a la classe dels ciclòstoms (els representants més coneguts són les llamprees) i la dels ostracoderms. Actualment constitueixen un grup a part, inclòs en la superclasse dels àgnats. Són vertebrats molt primitius i una de les seves característiques principals és l’absència de mandíbula.
La metamorfosi
Dels ous dels animals no sempre es forma directament un organisme semblant al que serà l’individu adult, un cop acabat el període de creixement i assolida la maduresa. En molts casos es forma una larva, notablement distinta de l’adult en què es convertirà directament o a través d’altres estadis larvals, diferents de l’inicial. El conjunt de transformacions que patirà l’animal en aquests casos s’anomena metamorfosi.
Hi ha molts grups animals que presenten metamorfosi: tots els invertebrats aquàtics i paràsits, els tunicats, alguns peixos, els insectes i els amfibis. Els rèptils, els ocells i els mamífers, per bé que, com és natural, pateixen variacions d’aspecte abans d’arribar a l’estat adult, no experimenten cap metamorfosi: són els anomenats animals de desenvolupament directe.
La metamorfosi és molt interessant en els insectes i els amfibis. Les diferències entre les larves i els adults són de vegades molt marcades, sobretot quan els dos estadis de l’organisme viuen en un medi diferent (la larva a l’aigua i l’adult a la terra ferma) i no tenen els mateixos hàbits alimentaris. Les larves dels insectes mai no tenen ales (àpteres), de vegades són aquàtiques (com el mosquit) i en molts casos devoradores de fulles o d’altres substàncies vegetals (els cucs de la majoria de les papallones). En canvi, l’aparell bucal dels adults és sovint més apte per a xuclar líquids que per a mastegar (tanmateix, no manquen els adults devoradors i fins i tot carnívors).
Entre els insectes, la metamorfosi té lloc de maneres molt variades, amb diferències més o menys marcades entre la larva i l’adult i de vegades amb més d’un estadi larval diferent. És el cas dels holometàbols; els insectes lepidòpters en són un bon exemple. De l’ou de la papallona surt una larva vermiforme (eruga), força activa i voraç, que després d’una sèrie de mudes que li permeten augmentar les dimensions es transforma en crisàlide (de vegades dins un capoll). La crisàlide és una larva immòbil que no s’alimenta i a l’interior de la qual es produeixen canvis profunds que originaran l’insecte adult, amb ales molt variades, potes i probòscide bucal per a xuclar el líquid ensucrat (el nèctar) de què s’alimenta.
Pel que fa als amfibis anurs (les granotes i els gripaus), dels seus ous surt una larva que s’anomena renoc o capgròs. És aquàtica i, per tant, posseeix una cua per a nedar. No té extremitats i respira per mitjà de brànquies externes. El capgròs s’alimenta de substàncies vegetals que hi ha a l’aigua i té un tub digestiu molt llarg. En aquest cas la transformació en adult és gradual i consisteix fonamentalment en: la pèrdua de la cua, que és reabsorbida mentre creixen les potes; la reabsorció de les brànquies mentre els pulmons es desenvolupen i finalment esdevenen funcionals; l’escurçament de l’intestí mentre l’alimentació va esdevenint carnívora (els adults es nodreixen d’insectes); l’engruiximent del tegument per a passar a viure fora de l’aigua, i el perfeccionament de l’ull i de l’orella interna (es forma el timpà).
La reproducció
La finalitat de tot ésser viu és preservar l’espècie a la qual pertany. La condició essencial per a assolir aquest objectiu és la producció contínua d’individus nous amb les característiques típiques del grup. Els organismes inverteixen temps i energia, es comporten d’una manera complexa i s’arrisquen fins al punt de deixar-hi la pell per reproduir-se i dur a terme aquesta missió: assegurar la continuïtat de l’espècie (vegeu també "Societats i comportaments animals"). Hi ha dos tipus fonamentals de reproducció: la sexual i l’asexual. En el món animal el tipus de reproducció més freqüent és la sexual, en què un mascle i una femella "col·laboren" per generar un nou organisme. Tanmateix, la reproducció asexual també hi té certa difusió.
Reproducció asexual
En la reproducció asexual, el fill té un sol progenitor i es desenvolupa a partir d’un fragment, més o menys gros, que prové del cos d’aquest progenitor. En el cas dels organismes unicel·lulars que es reprodueixen per escissió, és a dir per subdivisió en dues cèl·lules filles més o menys iguals, el preu que ha de pagar el progenitor és elevadíssim, ja que deixa d’existir com a individu. És el mateix destí d’alguns celenterats i platihelmints que tenen el mateix tipus de reproducció. L’individu adult es divideix en dos o més fragments de mida aproximadament igual i no queda res de l’individu original. Després, cada fill creix i regenera les parts que li manquen fins que esdevé un individu complet.
Però el resultat de la reproducció asexual no sempre és tan dramàtic per al progenitor. En molts casos (els protozous ciliats, les esponges i els pòlips), l’individu mare sobreviu tranquillament perquè el fill, molt més petit, s’origina a partir d’una zona ben delimitada del seu cos. En aquests casos es parla de gemmació. Els fills es poden separar completament de la mare i ser autònoms, o bé quedar-s’hi units i originar una colònia (en són un exemple les esponges i els coralls, com hem descrit més amunt).
Un cas particular de reproducció és el de la reproducció a partir d’ous no fecundats. En aquest cas, com en la reproducció asexual, només participa un sol progenitor. És la partenogènesi, que s’observa entre alguns cucs, crustacis i insectes (sobretot himenòpters i coleòpters). Consisteix en el desenvolupament d’un ou que no ha estat fecundat pel mascle. Ara bé, hi ha diferenciació dels dos sexes i les mateixes femelles que es reprodueixen per partenogènesi poden reproduir-se sexualment per fecundació. Sembla que aquesta alternança té relació amb les situacions ambientals. En condicions favorables, com a l’estiu, es desenvolupen ous partenogenètics, mentre que en condicions més difícils, per exemple a l’hivern, se’n desenvolupen de fecundats. Això s’explica pel fet que, en una situació favorable, una reproducció que no depengui de la trobada dels dos membres de la parella permet una difusió dels individus ràpida i segura. En aquest període els mascles poden resultar del tot absents. En una situació ambiental desfavorable, en canvi, resulta més avantatjós per a la conservació de l’espècie la presència d’individus originats per fecundació i, per tant, amb combinacions de caràcters diferents. En aquest cas també serà més probable que es desenvolupin característiques favorables per a l’adaptació. Es pot donar la mateixa justificació pel que fa a les alternances de reproducció sexual i asexual.
En altres casos, la partenogènesi representa un mecanisme de determinació del sexe. Les abelles, per exemple, dipositen els ous fecundats a les cel·les previstes per a la reina o per a les obreres (durant el vol nupcial la reina ha estat fecundada i conserva els espermatozoides en un sac connectat a l’oviducte) i d’aquests ous sortiran femelles, mentre que els ous que no estan fecundats són col·locats en unes altres cel·les, de les quals sortiran els abellots, és a dir els mascles.
Reproducció sexual
Perquè hi hagi reproducció sexual calen, com ja s’ha dit, dos individus. Consisteix en la fusió (fecundació) dels gàmetes òvul i espermatozoide en una única cèl·lula, a partir de la qual naixerà un nou organisme. El fet que la reproducció més estesa en el món animal sigui la sexual pot ser fàcilment entès si es té en compte que, des del punt de vista evolutiu, representa una sòlida garantia de la conservació de l’espècie, a diferència de la reproducció asexual, en què els fills són genèticament idèntics als progenitors. L’individu que s’origina a partir de la fusió de les cèl·lules germinals de dos organismes diferents resulta òbviament diferent, tant dels pares (ja que és una barreja dels caràcters hereditaris de tots dos) com dels germans, atès que la barreja és casual.
D’aquesta manera, contínuament s’originen individus amb característiques originals respecte dels progenitors i amb combinacions de caràcters molt diverses, la qual cosa permet noves possibilitats d’adaptació al medi i té com a resultat una probabilitat més gran de supervivència per a l’espècie, fins i tot en condicions ambientals particularment desfavorables. Els protozous ja presenten reproducció sexual, amb modalitats molt variades. En alguns casos, per exemple en els flagel·lats, fins i tot els individus sencers es comporten com a cèl·lules germinals i, en fecundar-se, es fonen completament. En altres casos (els foraminífers i els esporozous) es formen petites cèl·lules germinals amb característiques que ja són habituals en els gàmetes d’animals més evolucionats. L’element femení és més gros i és ple de citoplasma immòbil, mentre que el masculí és petit i mòbil. Entre els ciliats, en canvi, dos individus es poden unir en la regió de la boca durant un curt període de temps i a través d’aquesta connexió citoplasmàtica intercanvien material genètic (conjugació).
Els animals pluricel·lulars tenen uns òrgans específics, les gònades, encarregats de produir els gàmetes. En general, cada individu té només un tipus de gònades i pertany a un sexe ben definit: és mascle si té testicles, i femella si disposa d’ovari.
Hi ha casos, però, en què un individu posseeix ambdós òrgans. Aleshores es parla d’animals hermafrodites; és el cas dels cucs plans, els anèl·lids i alguns tipus de mol·luscs. Aquest fenomen, rar entre els vertebrats, s’observa per exemple en alguns peixos. Tot i això, encara que tinguin testicles i ovaris, en general no es produeix autofecundació, sinó que l’aparell masculí d’un individu fecunda el femení d’un altre, constituint una mena de fecundació encreuada.
Mascles i femelles: les diferències externes
En general els animals de sexes separats, que són la immensa majoria, han desenvolupat característiques físiques com la forma, el color i les dimensions que distingeixen clarament els mascles de les femelles. Es parla aleshores de dimorfisme sexual.
En molts casos (els mol·luscs, els insectes, els peixos, els amfibis i els rèptils) les femelles són més grosses que els mascles. Hi ha situacions extremes en què els mascles són veritables "miniatures", com és el cas del cuc marí Bonellia viridis (vegeu "Casos particulars de determinació del sexe").
Un cas extrem, i no tan sols de dimorfisme sexual, és el d’un peix abissal, l’Edriolychnus: el mascle, que té gairebé tots els òrgans atrofiats, menys els testicles, s’aferra a la femella, que és deu vegades més gran, i esdevé pràcticament un paràsit de la seva companya. Fins i tot hi arriba a connectar el seu sistema circulatori.
Tot i que entre els mamífers i els ocells és generalment el mascle qui té la supremacia quant a les dimensions, si s’observa el món animal en conjunt es constata que hi ha un clar predomini d’espècies en què la femella és més grossa que el mascle. Aquesta afirmació es pot justificar fàcilment si es té en compte que tres quartes parts de les espècies animals pertanyen a la classe dels insectes, que és on el mascle sol ser més gràcil que la seva companya.
Però, també entre els insectes, els mascles gaudeixen gairebé sempre de més mobilitat (sovint les ales de les femelles són reduïdes) i tenen els òrgans sensorials més aguts. Aquestes característiques probablement es relacionen amb el fet que la recerca activa i el festeig de la parella solen ser tasques masculines.
Així, en els mascles les antenes sovint són més llargues i amb més cèl·lules sensitives (el borinot mascle en té 50 000 i la femella 10 000) i els ulls solen tenir més agudesa visual. A més, concretament entre els ocells i els peixos, els mascles són més bonics, amb un plomatge més variat i un aspecte general més vistós que els serveix per a seduir la femella i impressionar els seus rivals.
La primera condició perquè hi pugui haver fecundació és que es trobin un mascle i una femella de la mateixa espècie, i que siguin sexualment madurs, és a dir, capaços de reproduir-se. Amb aquesta finalitat, moltes espècies animals han elaborat senyals específics destinats a orientar un dels possibles companys envers l’altre, i viceversa. Són senyals de la més diversa mena i de vegades curiosos, característics de cada espècie. D’aquest tema se’n parla àmpliament a "Societats i comportaments animals".
Casos particulars de determinació del sexe
En la majoria dels casos, el sexe d’un individu depèn de la dotació genètica que té i per tant es determina en el moment de la fecundació (vegeu "Les lleis de la genètica"), però en moltes ocasions la determinació del sexe depèn de factors diversos. En certs animals, a més, pot ser que el sexe no sigui el mateix al llarg de la vida.
En alguns celenterats, anèl·lids, crustacis i gastròpodes, el sexe és determinat per factors externs, i això pot passar en qualsevol moment del desenvolupament de l’individu. Un exemple clàssic és el cuc marí bonèl·lia (Bonellia viridis). Els mascles són molt petits (d’1 a 2 mm) i viuen sobre la probòscide de la femella, que és molt més grossa (de fins a un metre, probòscide inclosa). Les larves d’aquest animal no són sexualment definides; si en nedar topen amb una femella s’instal·len damunt seu i esdevenen mascles per l’acció d’una hormona emesa per la companya i, pel que sembla, d’alguns ions "masculinitzants" presents en l’aigua de mar. En canvi, si no troben cap femella, aleshores esdevenen femelles.
Igualment, en les tortugues, i sembla que també en algunes sargantanes i en el caiman del Mississipí, el sexe és determinat pel medi, i en particular per la temperatura d’incubació dels ous: entre 26°C i 28°C neixen mascles, entre 32°C i 34°C neixen femelles, i a 30°C el 64% són femelles i el 36% mascles.
També hi ha animals que al llarg de la seva vida canvien de sexe: són hermafrodites. La primera part de la seva vida són femelles i després mascles, o viceversa. L’ostra és un cas a part, perquè canvia de sexe diverses vegades al llarg de la seva vida.
La fecundació interna
En la fecundació interna els òvuls es disposen a l’interior del sistema reproductor de la femella, i és aquí on els espermatozoides aniran a trobar-los i els fecundaran. En general, els espermatozoides emesos pel mascle estan barrejats amb un líquid que els ajuda a conservar-se vius i eficaços. Altres vegades, però, són en una mena de saquets anomenats espermatòfors.
Els espermatòfors s’insereixen en el cos de la femella de maneres molt diverses. En alguns insectes és la femella qui se n’encarrega. Hi ha uns poliquets on aquesta penetració és més aviat cruel: el mascle porta els espermatòfors als apèndixs posteriors i neda al voltant de la femella, que l’intercepta i, sense gaires miraments, li arrenca els apèndixs i els engoleix. Igualment, hi ha cucs plans on la introducció dels espermatòfors és més aviat decidida. Aquesta vegada són els mascles que s’encarreguen d’inserir-los, però ho fan en un punt qualsevol del cos de la femella.
Els escorpins són més formals: s’aferren l’un a l’altre i fan una mena de dansa nupcial durant la qual el mascle arrossega la femella fent-la passar per damunt dels espermatòfors, que va deixant anar a poc a poc.
Els mascles de molts cefalòpodes, en canvi, tenen un òrgan especial destinat a la inseminació: un braç copulador, recorregut per un canal, que s’insereix en la cavitat del mantell de la femella. Per mitjà de contraccions, els espermatòfors llisquen fins a l’obertura genital de la femella.
En tots aquests casos es tracta de fecundació interna, que es defineix com a indirecta perquè els espermatozoides resten un temps a l’exterior dels òrgans genitals. En canvi, en la fecundació directa, que és la fecundació interna més estesa en el món animal, els espermatozoides passen de l’aparell reproductor masculí al femení d’una forma immediata, bé mitjançant el simple acostament de les obertures genitals, com en els ocells, bé per un òrgan copulador com el penis, present en molts animals (els cucs plans, els gastròpodes, els insectes, els mamífers i els selacis). És un òrgan tubular recorregut pel canal deferent del líquid seminal que, introduït en l’aparell femení, diposita els espermatozoides sense dispersar-los.
La fecundació externa
Entre els animals que viuen a l’aigua (els celenterats, els mol·luscs, els peixos i els amfibis) és molt estesa una modalitat de fecundació que té lloc a l’exterior del cos de la femella. Tant el mascle com la femella posen a l’aigua un gran nombre d’espermatozoides i d’òvuls respectivament; atrets per substàncies químiques determinades, els espermatozoides fecundaran els òvuls.
Tot i que la fecundació externa és un procés que genera una despesa i dispersió enormes, ja que s’han de produir molts òvuls per a tenir certa seguretat que algun en sigui fecundat, la trobada entre els dos gàmetes no és un fet casual. Les maniobres de festeig i els rituals d’apariament, que són especialment difosos entre els animals aquàtics, també tenen la finalitat —un cop els dos membres de la parella s’han reconegut gràcies als senyals convencionals de l’espècie— de provocar l’emissió d’òvuls i espermatozoides al mateix lloc i en el mateix moment.
La protecció de l'embrió
Un cop produïda la fecundació, el problema és nodrir i protegir l’embrió fins que sigui completament madur. Els embrions dels mamífers placentaris es desenvolupen a l’interior del cos matern, dins un òrgan específic, apropiat per a aquesta funció, que s’anomena úter, i es nodreixen a través de la placenta. Els marsupials són un cas a part ja que no formen la placenta, i els petits, un cop consumides les substàncies nutritives de l’ou, han de ser parits en estat encara embrionari. Aleshores s’acaben de desenvolupar a l’interior d’una bossa —el marsupi— situada al ventre de la mare, a l’interior de la qual hi ha les mamelles.
En altres animals, com els selacis (els taurons, etc.) i alguns tipus de rèptils, que viuen en condicions ambientals particularment difícils (al desert, a la muntanya), els ous romanen ben protegits a l’oviducte, que els assegura oxigen i humitat, fins que eclosionen.
En alguns peixos (per exemple, els cíclids), els ous són incubats a la boca de la mare i així queden protegits dels predadors. Entre els cavallets de mar, en canvi, és el mascle que porta els ous dins una bossa incubadora ventral. També entre els invertebrats hi ha casos en què l’embrió continua gaudint durant un temps de la protecció que li ofereix el cos matern. En moltes actínies, les anomenades "anemones de mar", els petits completen el seu desenvolupament dins la cavitat gastrovascular materna.
La vagina de la mosca de la carn presenta una bossa incubadora on té lloc el creixement de l’embrió fins a l’estadi larval. Les femelles dels branquiòpodes, una espècie de crustacis, protegeixen els ous i els embrions en una bossa dorsal. Igualment, molts cucs troben refugi durant un temps a l’interior dels òrgans genitals materns.
L'ou
Hi ha molts casos en què l’ou es pon a l’exterior i l’embrió completa el seu desenvolupament separat del cos de la mare. El medi aquàtic és un ambient favorable, perquè l’aigua protegeix l’ou dels canvis forts de temperatura i de la deshidratació. Així, els ous postos a l’aigua no requereixen un revestiment especial, perquè no corren gaire perill (el problema dels predadors és d’una altra mena), i també poden ser fecundats un cop postos (fecundació externa). Els ous dels rèptils i els ocells, en canvi, han de tenir una closca protectora gruixuda, secretada per les glàndules que entapissen l’oviducte (conducte que va de les oueres a l’exterior). Això implica la fecundació interna, ja que l’espermatozoide ha de poder arribar a l’òvul abans que sigui embolcallat pels involucres protectors. Òbviament, la fecundació interna es dóna en els animals vivípars que no ponen els ous, sinó que aquests es conserven a l’interior del cos matern, on es desenvolupen, com és típic dels mamífers (llevat dels monotremes, com l’ornitorinc).
La grandària dels ous varia molt d’una espècie a una altra i no té relació amb les dimensions dels animals, sinó amb el tipus de desenvolupament embrionari. Entre els mamífers, els ous són molt petits perquè es desenvolupen a l’úter i, com ja s’ha dit, les substàncies nutritives els arriben del cos matern a través de la placenta. En aquells animals en què el desenvolupament de l’embrió té lloc d’una manera totalment independent de l’organisme matern, els ous, que es ponen a l’exterior, són molt més grossos, perquè han de contenir reserves alimentàries suficients per tal que la cria maduri completament.
El sistema nerviós
Si en un animal la freqüència dels batecs del cor fos casual, els músculs es contraguessin desordenadament, la sang fos distribuïda de manera irregular i no arribessin sensacions del medi extern, el resultat seria la mort. És fonamental que, en organismes complexos com els animals, hi hagi una coordinació precisa i un control constant de totes les funcions vitals, de la digestió al moviment, passant per la respiració i la reproducció, la circulació i l’excreció i els ritmes del son i la vigília. De la mateixa manera, per a sobreviure és indispensable el contacte i la interacció amb el món exterior, percebre estímuls i rebre informació, i poder generar les respostes apropiades.
Funcions del sistema nerviós
Totes aquestes tasques que acabem d’enumerar són dutes a terme pel sistema nerviós, un complex sistema de cèl·lules i fibres que, en molts casos, és ajudat per l’aparell endocrí. De vegades les plantes també presenten respostes als estímuls del medi, lluminosos, químics o de contacte. Però en general aquestes reaccions depenen de la circulació de certes substàncies i, per tant, són infinitament més lentes que els estímuls nerviosos (vegeu "Les plantes del nostre entorn").
El fet que els animals tinguin sistema nerviós i les plantes no, està probablement relacionat amb el moviment. A les plantes no els cal desplaçar-se per a obtenir l’aliment, només necessiten un sistema radical eficaç i certa superfície foliar. Els animals, en canvi, es mouen per satisfer les seves exigències vitals i necessiten gran quantitat d’informació del món que els envolta, una xarxa de comunicació interna rapidíssima i un sistema que ho controli i ho coordini tot.
Els sistemes nerviosos més simples
Ja en organismes unicel·lulars es troben indicis de sistema nerviós. D’altra banda, quan es parla d’organismes capaços de moure’s, d’aconseguir l’aliment d’una manera activa, de protegir-se de les situacions ambientals difícils, automàticament se suposa que hi ha d’haver alguna mena de sistema que coordina tot això.
La primera estructura nerviosa realment organitzada és visible en els protozous paramecis. Es tracta d’una xarxa de fibril·les situada immediatament al dessota de la membrana cel·lular, que controla directament el moviment dels cilis. Si es trenca es perd la coordinació del moviment i l’animal comença a nedar de manera desordenada i poc eficaç.
Aquests organismes demostren, a més, que capten les sensacions del medi i que el seu comportament està directament relacionat amb els estímuls externs. Si la temperatura és massa alta o massa baixa, o si és un medi massa àcid o amb massa oxigen, es retreuen, s’allunyen i canvien de direcció, i no s’aturen fins que troben condicions de vida acceptables.
Els organismes pluricel·lulars disposen de sistemes, més o menys desenvolupats, formats d’autèntiques cèl·lules nervioses. Són cèl·lules especialitzades amb llargues prolongacions a través de les quals viatgen els estímuls, que d’aquesta manera són transmesos d’una part del cos a una altra. Els celenterats, per exemple, presenten una xarxa nerviosa que coordina moviments simples relacionats amb la locomoció i la captura de la presa. Aquesta xarxa és composta per cèl·lules distribuïdes uniformement per tot el cos (són lleugerament més compactes al voltant de la boca). En aquests animals l’aplicació d’un estímul en un punt determinat té una difusió indiferenciada: es propaga en totes direccions, i s’afebleix a mesura que s’allunya.
La centralització del sistema
Els cucs plans i cilíndrics ja presenten un canvi important: la primera centralització del sistema nerviós. Al cap apareixen dues grans agrupacions de cèl·lules nervioses, els ganglis, a sobre i a sota del tub digestiu. A més, tot el cos és recorregut per dos cordons que es connecten per fibres transversals, com una escala de corda.
Aquesta tendència a situar l’agrupació principal de cèl·lules al cap i constituir un centre nerviós ben definit és més pronunciada en els anèl·lids i els artròpodes. En els anèl·lids hi ha un parell de ganglis cerebrals: un de superior, situat sobre l’esòfag, que fa la funció de control i es considera el cervell, per bé que la seva estructura encara és uniforme i sense centres específics; i un d’inferior doble, on va a parar la cadena de ganglis que recorre verticalment el cos de l’animal. Cada segment és innervat per un parell de ganglis i té, per tant, certa autonomia.
La cadena ganglionar també és responsable del moviment: organitza les contraccions musculars en ones que recorren tot el cos i per mitjà de les quals l’animal es mou. Però del control global del moviment se n’encarrega el cervell. Si aquest és danyat o inactivat d’alguna manera, l’animal es comença a moure sense parar.
La diferenciació del cervell
Entre els poliquets, els anèl·lids marins per excel·lència, predadors amb ulls i antenes, el cervell comença a presentar certa diferenciació. Les cèl·lules cerebrals se subdivideixen en grups amb funcions específiques i es comencen a distingir zones ben caracteritzades. Així, per exemple, el moviment de les antenes és controlat únicament per una àrea ben definida del cervell i no per tot l’òrgan sencer.
El centre nerviós dels insectes també se situa al cap, per sobre de l’esòfag, i és un gangli molt gros, format per milers de cèl·lules. És subdividit en zones molt precises, cadascuna de les quals controla i coordina una activitat. Es tracta d’un veritable cervell amb funcions complexes. D’altra banda, el comportament d’aquests animals és notablement complicat; només cal pensar en les complexes relacions que estableixen els insectes socials, com les abelles i les formigues, i en la seva capacitat de comunicació. Tot això implica la presència d’un centre nerviós força evolucionat. El gangli cerebral inferior, més simple, controla els moviments de l’aparell bucal tant pel que fa a la masticació com a la defensa. És on va a parar la cadena ganglionar ventral que, en aquest cas, també recorre tot el cos. Aquests ganglis tenen molta importància en els moviments reflexos i coordinen els moviments de les potes i de les ales.
El sistema nerviós dels cefalòpodes també es caracteritza per una gran complexitat i una centralització important. El cervell dels pops probablement és més evolucionat que el de cap altre grup, excepte els vertebrats. En els cefalòpodes s’ha comprovat la fusió dels ganglis cerebrals i viscerals, que ha originat un cervell gros situat al voltant de la faringe i circumdat per una estructura cartilaginosa de protecció, una caixa cranial rudimentària.
En aquest cas, l’estructura del cervell tampoc no es diferencia, però s’hi detecten diversos centres que controlen funcions diferents de complexitat variable. Per exemple a la part anterior hi ha un centre responsable de la capacitat de mossegar, mentre que més enrere hi ha una zona que regula el moviment dels tentacles per a la captura de la presa. Els estímuls d’aquests dos centres arriben després a una zona superior del cervell que els integra per tal d’obtenir un moviment de presa dels aliments ben sincronitzat. El desenvolupament excepcional d’aquest cervell es correspon a la complexitat de les funcions que duu a terme, un exemple de les quals és l’alimentació. Una altra funció particularment evolucionada i que requereix una regulació nerviosa sofisticada és la visió. Els ulls dels pops tenen una estructura fins i tot comparable a la dels vertebrats.
El sistema nerviós dels vertebrats
En els vertebrats el sistema nerviós assoleix un grau de complexitat i especialització molt superior al de qualsevol altre grup. S’estructura segons un model comú a tots ells. Al cap, protegit per la caixa cranial, se situa l’encèfal, que es prolonga en la medul·la espinal, cordó nerviós que recorre per l’interior de la columna vertebral tota la llargada del cos de l’animal, però en posició dorsal i no ventral com en els animals més primitius. L’encèfal i la medul·la espinal representen el sistema nerviós central i són constituïts per neurones. Les neurones són cèl·lules nervioses de les quals, com s’ha dit, surten llargues prolongacions, els axons, que s’agrupen en feixos que formen els nervis, distribuïts per tot el cos de l’animal. Són comparables a cables al llarg dels quals viatgen els missatges i els senyals nerviosos en forma d’impulsos elèctrics, a gran velocitat (fins a 300 km/h), que posen en contacte el sistema nerviós central i el món exterior. Per mitjà dels nervis sensitius el centre pot rebre informació del medi, captada per les estructures especialitzades dels òrgans sensorials, com també per mitjà dels nervis motors, que transporten els senyals dirigits als músculs, i pot organitzar reaccions, respostes i comportaments.
Els nervis, exceptuant els del cap, són connectats al cervell per les cèl·lules nervioses de la medul·la espinal. Però aquesta no és solament una estació de trànsit per als estímuls que van i vénen del cervell, ja que també representa el centre on neixen i es propaguen els reflexos, respostes involuntàries a estímuls determinats.
L’encèfal representa la millor agrupació de cèl·lules nervioses. En tots els vertebrats s’hi poden distingir 5 zones: el cervell anterior (telencèfal), el cervell intermedi (diencèfal), el cervell mitjà (mesencèfal), el cerebel i la medul·la oblonga. Aquesta darrera és semblant a la medul·la espinal i és una zona de trànsit a totes les vies, sensorials i motrius, que uneixen aquesta al cervell anterior. A més, és un centre de funcions importants: la medul·la oblonga elabora els reflexos de la respiració, la masticació, la sudació i el vòmit.
El cerebel, en canvi, regula l’equilibri i la coordinació dels moviments, i rep estímuls de tots els músculs, del laberint (orella interna), dels ulls i del cervell. Amb la destrucció del cerebel, els animals perden la capacitat de moure’s amb normalitat, amb l’adequada coordinació muscular. El cerebel és més aviat petit en els peixos i els amfibis, i és cada vegada més gros en els rèptils, els ocells i els mamífers.
El mesencèfal dels vertebrats menys evolucionats és un centre visual al qual van a parar les fibres del nervi òptic i també les del nervi laberíntic; és important, doncs, perquè reuneix estímuls relatius a l’equilibri i estímuls visuals. En els peixos, que tenen l’escorça cerebral poc desenvolupada, és el centre d’associació més important i seu dels instints i els processos d’aprenentatge.
El diencèfal representa sobretot una zona d’encreuament dels estímuls procedents de l’escorça cerebral i els de les altres àrees cerebrals. Hi ha, per exemple, una zona per a l’encreuament dels estímuls olfactius amb el control dels moviments de la boca, important per a l’obtenció de l’aliment. La base del diencèfal, l’hipotàlem, regula funcions vegetatives importants i vitals: la respiració, la pressió sanguínia, el ritme de son i vigília, la sensació de gana i set, la temperatura.
Al llarg de l’evolució, el telencèfal dels organismes ha esdevingut la part més important del cervell. Externament apareix format per 2 hemisferis revestits per l’escorça cerebral, que és constituïda per un nombre elevadíssim de neurones (vegeu "L’escorça cerebral").
L’escorça cerebral
En els vertebrats més evolucionats, l’escorça cerebral és un centre de coordinació i d’integració d’importància fonamental. Conté les àrees on es registren i s’elaboren les sensacions i les percepcions, es confeccionen les respostes motores, s’organitza el comportament i es posen en marxa els processos d’aprenentatge (vegeu també "La regulació de l’organisme").
La capacitat major o menor de realitzar accions complexes, d’adaptar el propi comportament a les característiques del medi i de modificar-lo segons l’experiència, és a dir, d’aprendre, està directament relacionada amb el desenvolupament i la complexitat de l’escorça cerebral. Pujant en l’escala zoològica n’augmenta enormement l’extensió i, consegüentment, els solcs són més profunds i més complicats, per tal que, a igualtat de volum, pugui allotjar un nombre de cèl·lules molt més elevat. Paral·lelament, augmenta la capacitat d’aprenentatge i la ductilitat del comportament. Es passa de comportaments també complexos, però instintius, és a dir, programats genèticament i per tant rígids, a comportaments apresos i modificables a partir de l’experiència individual.
Alguns insectes, com les formigues, tenen una estructura social complexa i complicades relacions entre ells. Són capaços de comunicar-se, però no aprenen, i els seus comportaments són fixats i predeterminats.
Fa uns anys, a Anglaterra, algunes mallerengues van aconseguir una nova font d’aliment aprenent a foradar amb el bec el tap de les ampolles de llet que el repartidor deixava al carrer davant la porta de les cases. S’ha vist com els bernats pescaires pescaven fent servir una ploma d’esquer per atreure els peixos. Els simis antropomorfs, que tenen una escorça cerebral molt desenvolupada, han estat observats elaborant estratègies abstractes, utilitzant instruments construïts expressament amb una finalitat precisa (per exemple, branquillons amb les fulles arrencades per "pescar" els tèrmits del termiter) i actuant d’una manera generalitzada, és a dir, adonant-se que el mateix objecte pot tenir més d’una destinació: un bastó es pot fer servir per a abastar aliments inaccessibles o per a amenaçar un adversari. El major desenvolupament de l’escorça, doncs, es manifesta en la varietat d’usos que un animal pot donar a un objecte, en la capacitat d’aprofitar els elements nous del medi, i en la facultat de recombinar elements diferents i fer servir recursos nous.
Amb l’evolució de l’escorça, on cada vegada es concentren més funcions, augmenta la seva importància fins i tot pel que fa a la supervivència de l’animal. Així, mentre que la destrucció de l’escorça gairebé no té cap efecte en els peixos, perquè la tenen molt poc desenvolupada, és causa de problemes seriosos en els mamífers inferiors. Sense escorça, els simis poden sobreviure pocs dies, mentre que els humans moren immediatament.
La sensibilitat dels animals
Els animals, per a viure, han de ser conscients en tot moment de les condicions del medi on són i dels esdeveniments que s’hi produeixen, i a la vegada de la situació del propi cos, tant internament com en relació amb el medi. És una consciència construïda per una enorme quantitat d’informació captada i registrada per estructures especialitzades: els receptors i els òrgans sensorials, de complexitat i naturalesa molt variades. Hi ha receptors formats tan sols per una terminació nerviosa simple i òrgans sensorials extremament sofisticats.
No tots els estímuls que provenen del medi són registrats indiscriminadament, sinó que se seleccionen els que són fonamentals per a l’organisme. És sobretot així com s’obtenen les reaccions als estímuls químics, lumínics, mecànics, tèrmics i sonors.
Estímuls químics
Tots els animals capten, per bé que amb modalitats diferents, els estímuls de naturalesa química. És precisament la capacitat de detectar determinades substàncies dissoltes en l’aigua o suspeses en l’aire en molts casos allò que fa possible defensar-se dels predadors, reconèixer el propi territori i escollir la parella. Fins i tot entre els organismes unicel·lulars ja s’observa una sensibilitat d’aquest tipus, que provoca reaccions positives enfront de les substàncies nutritives i el rebuig enfront de les altres. Però tan sols els mol·luscs, els artròpodes i els vertebrats tenen cèl·lules sensorials específiques: els quimioreceptors. Les antenes i els apèndixs bucals dels crustacis (un animal de sensibilitat excepcional és el llamàntol, que té un sentit del gust milions de vegades més desenvolupat que els humans), la cavitat i la vora del mantell dels mol·luscs i els tentacles dels pops tenen quimioreceptors molt senzills. En canvi, els insectes i els vertebrats confien la sensibilitat als estímuls químics als veritables òrgans del gust i de l’olfacte.
Entre els insectes, el gust se sol localitzar a la boca, als apèndixs bucals i a la part anterior de les potes. Per exemple, si es toca la superfície inferior de la pota d’una papallona amb un pinzell sucat en aigua ensucrada, allargarà immediatament la probòscide. Els òrgans de l’olfacte se situen, en canvi, a les antenes (algunes papallones i alguna espècie de coleòpter també en tenen als apèndixs bucals) i tenen gran capacitat de percepció, semblant a la dels vertebrats.
Alguns grups d’animals perceben olors que els humans no poden captar o que perceben amb una intensitat infinitament inferior. Alguns insectes que s’alimenten de vegetals, per exemple, són molt sensibles a l’olor dels olis essencials de les plantes de què es nodreixen; d’altres, com els escarabats, fins i tot es comuniquen amb un codi d’olors. Algunes papallones nocturnes, lepidòpters, sembla que són capaces de percebre la substància que la femella fa servir com a reclam sexual a 11 km de distància.
Els vertebrats tenen més cèl·lules específiques sensibles al gust, generalment situades a la cavitat bucal i sobre la llengua. Alguns peixos amb les escates reduïdes tenen els receptors del gust escampats per tota la superfície del cos. Els vertebrats més evolucionats perceben, com els humans, quatre gustos fonamentals: el dolç, el salat, l’àcid i l’amarg. N’hi ha d’altres, en canvi, als quals manca la percepció d’alguns gustos i que presenten diferents llindars de sensibilitat. El gust dolç és l’únic que sempre es percep.
Entre els vertebrats, l’olfacte es localitza en la mucosa nasal. En sobresurten cèl·lules amb fins pèls sensorials que funcionen com a detectors: després de captar molècules en suspensió en l’aire, envien al cervell els missatges olfactius, que sovint són molt refinats.
Entre els mamífers hi ha molts grups que presenten una sensibilitat particularment desenvolupada; són els carnívors, els ungulats, els insectívors i els rosegadors. En aquests casos, el nombre de cèl·lules olfactives és molt elevat, de centenars de milions. El gos, per posar un exemple, en té uns 220 milions i gaudeix d’una sensibilitat elevadíssima per a les olors, fins a 1 milió de vegades la sensibilitat humana. Sembla que aquests animals en tenen prou amb una sola molècula per a obtenir l’estimulació i enviar el missatge al cervell.
Els rèptils tenen un òrgan olfactiu suplementari, l’òrgan de Jacobson, que funciona amb l’ajut de la llengua. La substància olorosa és captada per l’extrem de la llengua i conduïda al paladar, on s’allotja aquest òrgan. Quan una serp avança agitant la llengua, no té en realitat intencions amenaçadores; simplement tracta d’olorar-ho tot per a trobar indicis del possible pas d’una altra serp o d’una presa.
La vista
Pràcticament tots els animals tenen el sentit de la vista. Si la percepció de veritables imatges requereix òrgans complexos com els ulls, presents solament en els mol·luscs, els artròpodes i els vertebrats, la capacitat de reaccionar a la llum és present, això no obstant, en els organismes més simples. Així, ja alguns protozous presenten una estructura sensible a la llum, la taca ocular, i altres animals com els celenterats, les larves d’insectes i els anèl·lids mostren cèl·lules fotoreceptores escampades per tota l’epidermis. En aquests casos la percepció de la llum desencadena generalment reaccions de por o de fugida. Les cèl·lules sensitives, esteses per tot el cos, no s’il·luminen alhora ni amb la mateixa intensitat, la qual cosa permet a l’altre organisme reconèixer la direcció d’on prové la llum i allunyar-se’n.
Alguns celenterats, els cucs, els gastròpodes i les estrelles de mar tenen les cèl·lules fotosensibles agrupades i constituint el primer ull rudimentari, anomenat "de fosseta". La llum és retinguda per un pigment contingut en algunes cèl·lules, i hi ha una capa de substància transparent que funciona com una lent. Tot i això, la percepció d’imatges encara no és possible i tan sols poden captar la direcció i el moviment de l’estímul lumínic.
Els ulls en forma de calze, presents per exemple entre els poliquets marins, són òrgans més perfeccionats, on ja és possible la visió, per bé que rudimentària, de les formes. Aquests ulls tenen a la part anterior un forat molt petit i, tot i l’absència d’un sistema de refracció, les figures són projectades invertides a la retina, que és formada per les cèl·lules sensorials. L’especialització extrema d’aquesta mena d’ulls és representada pels ulls amb cristal·lí, estructura de lent que recull i dirigeix els raigs de llum i garanteix la percepció d’una imatge nítida en qualsevol condició. Aquests ulls són típics dels vertebrats i els cefalòpodes i els fan capaços de captar perfectament les formes i per tant de veure-hi en el sentit més complet del terme. Són ulls tan perfeccionats que es poden adaptar ràpidament als canvis d’intensitat de la llum i variar la distància d’enfocament bo i mantenint la mateixa nitidesa de la imatge.
L’estructura d’aquests òrgans presenta certa uniformitat en les diferents espècies, més enllà d’algunes adaptacions concretes. Són òrgans formats per: un revestiment sòlid (l’escleròtica), una capa transparent (la còrnia), l’iris, el cristal·lí, el cos vitri mucilaginós, la retina i un teixit pigmentat (la coroide).
En tots els vertebrats amb bon poder visual, la retina presenta una part central on les cèl·lules sensibles són molt fines. Així, la imatge produïda, constituïda per un nombre més gran de punts d’estimulació, resulta particularment nítida. En canvi, no tots els vertebrats gaudeixen de visió en color. Mentre que els simis superiors poden distingir tots els colors, molts dels ocells i els peixos tan sols en veuen uns quants i els gossos, els cavalls, els cérvols i en general tots els animals nocturns solament hi veuen en blanc i negre (els colors són percebuts com matisos del gris).
Els amfibis tenen una sensibilitat especial per al moviment. Copsen amb claredat les coses que es belluguen, però tenen una visió molt poc detallada; la construcció de les imatges és resultat únicament de la contraposició de zones d’ombra i de llum.
Una altra mena d’ull que permet la visió d’imatges és l’ull compost que posseeixen la majoria dels artròpodes i alguns mol·luscs. Aquest ull és format per moltíssims elements situats un al costat de l’altre, els ommatidis, cadascun dels quals és un ull petitíssim, proveït de còrnia, cristal·lí i retina. Així, la imatge obtinguda és la suma o la superposició de milers de fragments, cadascun dels quals correspon a un ommatidi.
En els casos en què la imatge es forma per suma, cada ommatidi és protegit òpticament per uns pigments, per tal que només sigui estimulat pels raigs de llum directes. La imatge final resulta força nítida. En canvi, els animals nocturns tenen en general ulls compostos per superposició. En aquest cas, els ommatidis no són aïllats òpticament i també reben els raigs de llum reflectits pels cristal·lins dels ommatidis propers. D’aquesta manera s’aconsegueix l’aprofitament òptim de la llum de poca intensitat, però la imatge final resulta més aviat confusa. Molts insectes tenen tots dos tipus d’ulls. Les libèl·lules, per exemple, en un primer moment enfoquen la presa amb els ommatidis que funcionen per superposició, molt lluminosos, i quan la tenen a prop enfoquen la imatge amb els ommatidis per aposició, situats a més profunditat. Respecte dels altres tipus d’ulls, els dels artròpodes posseeixen una característica específica relacionada amb la capacitat de volar: aconsegueixen percebre com a imatges aïllades els estímuls de successió molt ràpida. En l’home, quan les imatges immòbils se succeeixen a un ritme de 18-25 per segon, es fonen. El cinema es basa en aquest fenomen: en una pel·lícula, una sèrie d’imatges estàtiques apareixen en moviment perquè es passen davant els ulls dels espectadors a una velocitat superior a la "freqüència de fusió".
En general, els insectes diürns són capaços de distingir els colors; a més, diversos himenòpters (les abelles i les vespes) i dípters (els mosquits) perceben les radiacions ultraviolades, que són invisibles per als humans.
Estímuls mecànics
El tacte és un altre sentit que presenten tots els animals, si bé amb modalitats diferents. Sovint els receptors es distribueixen per tota la superfície corporal, però són més nombrosos a les parts més exposades a l’aire lliure. Entre els invertebrats solen ser agrupacions de cèl·lules sensitives associades a estructures mòbils com cilis o pèls flexibles, tan sensibles que també perceben les vibracions de l’aire. En molts casos, els sistemes tàctils d’aquest tipus actuen també com a propioreceptors, és a dir, informen el sistema nerviós central de la posició recíproca de les diverses parts del cos. En molts insectes, per exemple, es troben a les articulacions de les potes, a l’extrem posterior del cap, entre el tòrax i l’abdomen, i mantenen sota control la posició d’aquestes parts del cos.
A l’epidermis dels vertebrats tant hi ha terminacions nervioses, lliures o associades a cèl·lules especialitzades, com receptors tàctils més organitzats. Molts carnívors tenen al musell i a l’esquena pèls llargs, connectats a les fibres nervioses, que exerceixen una funció tàctil.
En general, els animals responen als estímuls tàctils i a l’absència d’estímuls amb comportaments característics. La reacció immediata a un estímul tàctil sol ser defensiva, de fugida, o agressiva (mossegades, cops, picades) i, fins i tot, en molts insectes és la simulació de la mort. En alguns casos depèn de la intensitat de l’estímul; així, un lleuger fregament fa que l’animal s’allunyi, però el mateix contacte, si és més intens, pot desencadenar una violenta reacció defensiva.
L’absència d’estímuls determina generalment un comportament particular; l’allargament de les extremitats, la flexió del cos i les potes són els típics moviments que es fan en la caiguda o per preparar-se a "aterrar". Evidentment, l’absència de sensacions tàctils provoca la sensació del buit i la caiguda. Molts crustacis, en canvi, quan no detecten la presència d’objectes sòlids, amb els receptors de les potes comencen automàticament a efectuar moviments de natació.
Entre els peixos i els amfibis també hi ha òrgans tàctils especials situats al cap i als costats del cos. Són grups de cèl·lules proveïdes de bastonets sensorials que sobresurten i que no solament reaccionen enfront dels contactes directes, sinó que també són capaços de percebre les variacions de la pressió de l’aigua i localitzar d’aquesta manera els possibles obstacles.
Els peixos tenen, a més, un altre òrgan, anomenat "línia lateral", que els permet fer un càlcul de les distàncies. En els casos més simples està enfonsat en llargues fenedures laterals o bé en canals que comuniquen amb l’exterior per mitjà de porus situats darrere les escates. Els receptors d’aquest òrgan són estimulats pel moviment de l’aigua i per les variacions de pressió.
Estímuls tèrmics
La temperatura ambiental és un element molt important per a la vida de qualsevol animal. Un fort escalfament (superior als 45°C) o un fred intens representen un perill del qual els organismes s’han de protegir. En totes les espècies hi ha un interval de temperatura òptima dins el qual els processos vitals es produeixen amb la màxima eficàcia i la mínima despesa d’energia.
Per tant, tots els animals han de ser prou sensibles per a poder controlar en tot moment les condicions tèrmiques de l’ambient. Aquesta sensibilitat sol anar a càrrec de les terminacions nervioses lliures de l’epidermis. Els insectes tenen uns pèls especials disposats a l’últim tram de les antenes i als apèndixs maxil·lars que funcionen com a termoreceptors.
Les serps de cascavell tenen al cap una estructura sensorial específica que és sensible a les radiacions infraroges emeses pels éssers vius. Aquests animals són caçadors nocturns i probablement aconsegueixen localitzar la presa en la foscor gràcies a la calor que desprèn.
Els vertebrats presenten dos tipus de receptors: un de sensible al fred, situat quasi a la superfície, i un altre de sensible a la calor, a més profunditat. Per sobre de 40°C els receptors del fred deixen de funcionar, però si les temperatures són molt altes tornen a ser estimulats. Els receptors per a la calor funcionen fins a 10°C.
Estímuls sonors
Només els artròpodes i els vertebrats, animals que en general també produeixen sons, tenen veritables òrgans de l’oïda. Els organismes menys evolucionats capten els estímuls acústics a través de receptors semblants als tàctils. En els insectes, sovint els receptors d’aquest tipus també estan associats als receptors auditius. Els mosquits, per exemple, tenen a la base de les antenes un òrgan que tant reacciona a les sensacions tàctils com a les ones sonores.
Altres insectes, com les llagostes, les cigales i els grills, posseeixen òrgans especials per a percebre els sons. Són òrgans dotats d’una estructura semblant a un timpà. Es tracta de la paret d’una cavitat traqueal que comença a vibrar gràcies a les ones sonores i transmet l’estímul a les cèl·lules sensorials, les quals envien l’estímul nerviós. En alguns casos la sensibilitat auditiva és molt selectiva i està dirigida a interessos ben concrets; per exemple, en el mosquit Anopheles mascle la màxima sensibilitat auditiva correspon a la freqüència del so que produeixen les ales de la femella quan baten.
Els insectes en general són sensibles als sons de freqüència més aviat alta, fins i tot als ultrasons (per sobre de 21 000 Hz) que els humans no poden sentir. Moltes papallones nocturnes reaccionen a aquestes freqüències fugint o fins i tot simulant la mort per protegir-se dels seus predadors principals, com els ratpenats, que precisament emeten ultrasons per localitzar-les.
Els vertebrats, i en particular els mamífers, tenen l’oïda molt desenvolupada i localitzada en un òrgan complex, l’orella. Les ones sonores fan vibrar la membrana del timpà, que, a través d’un conjunt d’ossets, transmet la vibració a l’orella interna, on és captada per les cèl·lules sensorials.
Els mamífers, que en comparació amb altres animals com els rèptils i els amfibis tenen l’orella interna a més profunditat dins el crani, posseeixen també orella externa, el pavelló auditiu, que condueix les ones sonores cap al canal auditiu. Alguns ocells com l’òliba se’n poden "construir" un redreçant les plomes al voltant de l’orella. El pavelló auditiu dels ungulats i els carnívors és molt mòbil i fins i tot es pot orientar en la direcció d’on prové el so, que en els vertebrats terrestres es calcula a partir de les diferències de temps i d’intensitat amb què les dues orelles el capten.
Mentre que per a un ésser humà el llindar auditiu màxim és de 20 000 Hz, molts altres vertebrats superen de molt aquest límit. Els ratpenats, per exemple, capten fins a 170 000 Hz i ells mateixos produeixen ultrasons per orientar-se en la foscor i localitzar la presa. Per fer-ho es basen en l’eco produït pels seus crits en topar contra un objecte sòlid que els reflecteix. Els ratpenats emeten en repòs uns 8 sons per segon, durant el vol uns 30 per segon, i 50 o 60 quan són a prop d’algun obstacle.
Sembla que les rates, els ratolins, els talps, les balenes i els ocells que viuen en grutes també es poden orientar de la mateixa manera.
Contràriament al que es podria pensar, els peixos també gaudeixen d’una oïda ben desenvolupada, per bé que, en general, no són capaços de reconèixer la direcció dels sons. En moltes espècies és la mateixa bufeta natatòria que funciona com a caixa de ressonància, bo i reforçant el so que arriba.
L'equilibri
En tots els animals pluricel·lulars hi ha òrgans destinats a controlar la direcció de l’eix corporal respecte de la força de gravetat i per tant l’equilibri del cos en l’espai.
En els invertebrats hi ha els estatocists, vesícules plenes de líquid en què una partícula mòbil de calcària, sorra o matèria orgànica pressiona els cilis d’un grup de cèl·lules sensorials de la paret. Quan l’animal canvia de posició, l’estatòlit (la partícula) es mou i estimula altres cèl·lules. Així, el sistema nerviós central s’assabenta que hi ha hagut un desplaçament. La resposta és donada en forma de moviments compensatoris de tipus reflex, que serveixen per a restablir l’equilibri. En animals que troben refugi en el fang i la sorra, com els cucs i els mol·luscs, els estatocists serveixen per a controlar la direcció de l’enfonsament.
Els insectes no tenen estatocists. Probablement són els propioreceptors, situats al cap, i entre el tòrax i l’abdomen, els que perceben la direcció de la força de gravetat sobretot durant el vol, quan el cap i l’abdomen queden molt encorbats respecte del tòrax, que porta les ales.
Algunes xinxes d’aigua, que habitualment s’enfilen per les herbes aquàtiques i han de ser conscients de la noció "sobre i sota", tenen un òrgan curiós: als costats de l’abdomen s’obren dues fenedures en les quals, segons la direcció del moviment, una bombolla d’aire estimula uns pèls sensorials situats a altures diferents.
Tots els òrgans d’equilibri dels vertebrats presenten certa uniformitat d’estructura. Hi ha estatòlits a la zona de la base del laberint (orella interna) a tots dos costats del cap. En la posició normal, els pèls sobre els quals recolza l’estatòlit són adaptats a la pressió exercida pels pèls, però si l’animal s’inclina els pèls s’encorben, cosa que també determina un augment de la tensió dels músculs, amb la finalitat de restablir la posició normal; si l’animal es tomba, aquests mateixos pèls pateixen una violenta tracció que provoca un estímul particularment fort. Al laberint hi ha, a més, els tres canals semicirculars que informen el cervell de la direcció i, sobretot, de les variacions del moviment del cos. La seva funció és percebre anticipadament quan està a punt de produir-se una pèrdua d’equilibri.
Aquests canals contenen limfa i estan disposats aproximadament en els tres plans de l’espai.
Quan comença un moviment, la limfa del canal en el pla del qual té lloc el moviment es mou per inèrcia, al començament en direcció oposada, i exerceix pressió sobre una vàlvula situada a l’extrem del canal. La vàlvula es desplaça i determina el plegament dels pèls sensorials i, per consegüent, l’estimulació de les cèl·lules nervioses que envien missatges al cerebel, on s’organitzen moviments compensatoris. Passa el mateix quan el moviment s’atura o hi ha un canvi brusc de direcció, fets que poden precedir, i de manera força immediata, la pèrdua de l’equilibri.
La respiració i la circulació
Tots els organismes consumeixen energia per a moure’s, créixer, reproduir-se i viure en general. Aquesta energia és subministrada per les substàncies nutritives. Les molècules que han restat disponibles per al procés de la digestió, a l’interior de les cèl·lules del cos animal, són sotmeses a complexes reaccions químiques que alliberen energia. En aquest procés es consumeix oxigen i es produeix diòxid de carboni. Els animals solen tenir estructures especials com les tràquees, les brànquies i els pulmons perquè l’oxigen necessari per a eliminar el CO2 arribi a les cèl·lules.
La respiració en els animals més simples
Hi ha animals que no tenen cap òrgan especialitzat per a la funció respiratòria. Es tracta d’organismes més aviat simples, amb un tipus de respiració semblant a la de les plantes. L’oxigen és absorbit i el diòxid de carboni és eliminat per difusió a través de la superfície corporal.
Atès que la distància que l’oxigen aconsegueix recórrer per difusió no és gaire superior a 1 mm, aquest mecanisme funciona en organismes molt petits, com els protozous, o en animals més grossos, però amb el cos cilíndric o en forma de sac i amb les parets molt primes. Així, les cèl·lules són a una distància molt curta del fluid que circula per la cavitat i transporta l’oxigen, que hi pot arribar fàcilment. És el cas dels celenterats (l’aigua que transporta matèria nutritiva i oxigen entra a la cavitat digestiva), alguns gastròpodes marins i els cucs plans i els cilíndrics. Aquests animals tenen cavitats internes per on passa un líquid amb una funció anàloga a la de la sang, transportant l’oxigen que circula per mitjà de contraccions de les parets.
Uns altres animals, molt més grossos, que presenten un tipus de respiració anàloga, per difusió a través de la superfície cutània, són els amfibis. A l’estiu, les granotes cobreixen gairebé una tercera part de les seves necessitats absorbint oxigen a través de la pell i eliminen de la mateixa manera una quantitat de diòxid de carboni sis vegades superior a la que passa a través dels pulmons. En canvi, en els animals més grossos, la simple difusió a través de la pell no és suficient per a assegurar el subministrament d’oxigen. Hi ha, a més, un altre obstacle que fa gairebé impossible aquest tipus de respiració: a mesura que els animals esdevenen més complexos, la pell s’especialitza cada vegada més com a embolcall protector i esdevé menys permeable. Els intercanvis respiratoris han de ser duts a terme per estructures especialitzades: bé per parts diferenciades de la superfície corporal, exclusivament destinades a aquesta funció, com les brànquies, o bé per estructures internes específiques com els pulmons i les tràquees dels insectes.
Respirar a l'aigua: les brànquies
Les brànquies més simples són zones de la superfície corporal en què la pell és particularment prima i permeable a l’oxigen dissolt a l’aigua i amb abundants vasos sanguinis. Per augmentar la superfície d’intercanvi, aquesta zona forma plecs o, en situacions extremes, fins i tot origina apèndixs externs. És el cas dels cucs marins, que viuen en galeries excavades a la sorra o en els fons fangosos, on l’oxigen és escàs i l’han de captar de l’aigua del damunt. També és el cas de certs anèl·lids coberts per un revestiment rígid i impermeable en forma d’estoig.
Aquestes brànquies, de gran superfície i que sobresurten del cos, són molt vulnerables i, en animals que menen una vida molt més moguda que la dels cucs dels fons marins, serien una limitació molt important. Així, els animals aquàtics més grossos i amb activitats més complexes presenten brànquies protegides en cavitats especials del cos i incorporen dispositius que fan que circuli aigua nova contínuament, de manera que els intercanvis respiratoris tinguin lloc sense interrupció.
En la majoria dels mol·luscs, les brànquies són a la cavitat del mantell; en els gastròpodes branquiats i els bivalves són a l’interior de la closca i l’aigua circula gràcies al moviment dels cilis situats tant a la paret interna de la cavitat del mantell com a les mateixes brànquies; en els pops i les sípies, en canvi, hi ha músculs actius per bombar l’aigua cap a les brànquies.
En els crustacis, les brànquies, en forma de plomall, se situen lateralment al tòrax, en una cavitat formada per l’exosquelet.
En els peixos apareixen als costats del cap i en general són constituïdes per làmines disposades radialment i recorregudes per un gran nombre de capil·lars sanguinis. Se situen en una cavitat recoberta per l’opercle, una placa rígida però mòbil, i comuniquen amb la faringe, immediatament darrere la boca. L’aigua entra per la boca i passa per les brànquies, on cedeix l’oxigen als capil·lars, i després surt per l’obertura de l’opercle, és a dir per les ganyes. Per tal que l’aigua segueixi la direcció adequada del flux, de la boca a les brànquies, i un cop ha passat no torni enrere, hi ha unes vàlvules especials, tant al nivell de la boca com de l’opercle. Aquest, a més, té músculs, i exerceix una enèrgica acció de bombatge.
En alguns peixos bastinals, com les rajades, els taurons i els peixos martell, és el mateix moviment que determina el corrent d’aigua a les brànquies. Per a assegurar-se una oxigenació adequada, aquests peixos han de nedar contínuament; aturar-se durant massa estona els podria causar la mort per asfíxia.
Tant en les brànquies dels peixos com en les d’alguns mol·luscs, el flux sanguini al llarg de la superfície branquial va en direcció contrària al flux d’aigua. Gràcies a aquest fet tan senzill s’aconsegueix un aprofitament òptim de l’oxigen dissolt a l’aigua, ja que el pas de l’oxigen és causat per la seva diferent concentració a l’aigua i a la sang (és a dir que es produeix segons gradient de pressió). En efecte, en aquest intercanvi "a contra corrent", la sang que arriba a les brànquies (amb poquíssim oxigen) i avança, cada cop amb més oxigen, troba primer l’aigua que ja ha cedit part del seu oxigen, i després més aigua amb una concentració d’oxigen cada cop més gran, fins al moment de deixar les brànquies, que és quan troba l’aigua amb la màxima oxigenació. Així, els peixos obtenen de l’aigua del 80 al 90% de l’oxigen, mentre que els mamífers el capten de l’aire només en una proporció del 20 al 25%. El percentatge d’oxigen que hi ha en l’aire és del 21%, mentre que en l’aigua oscil·la entre el 0,5 i l’1,5%. La quantitat depèn de la temperatura i la pressió; l’aigua té el màxim contingut d’oxigen a 4°C, contingut que es redueix sensiblement a mesura que augmenta la temperatura (a 25°C és la meitat). És per això que els peixos que necessiten molt oxigen, per exemple les truites, prefereixen les aigües fredes dels torrents de muntanya.
Respirar a l'aire
Intuïtivament es podria pensar que la respiració dels animals terrestres és més fàcil que la dels animals aquàtics perquè l’aire, com ja s’ha dit, és molt més ric en oxigen. En realitat això no és cert, perquè els animals terrestres han desenvolupat revestiments encara més gruixuts i menys permeables que fan impossible qualsevol forma de difusió directa d’oxigen i de diòxid de carboni a través de la pell i, sobretot, perquè la vida terrestre planteja un problema d’importància fonamental: l’evaporació.
Com que l’oxigen tan sols pot penetrar en les cèl·lules per osmosi, dissolt en solució, les superfícies a través de les quals tenen lloc els intercanvis respiratoris han de ser humides (de manera que aquest gas pugui "dissoldre’s" en l’aigua i penetrar a l’interior) i primes per a fer-los possibles. Així, l’evaporació comporta un perill molt greu per als animals terrestres, un perill que cal tenir sempre present.
Alguns animals, com per exemple els cucs cilíndrics i els cucs de terra, que respiren a través de la pell, han resolt el problema vivint exclusivament en llocs humits, cosa que els fa molt vulnerables a qualsevol canvi ambiental. Si la humitat disminueix o, fins i tot, resten exposats al sol, la pell s’asseca i els és més difícil respirar. En canvi, els vertebrats i els artròpodes terrestres com els insectes han desenvolupat sistemes respiratoris que els protegeixen d’aquest perill i els permeten viure sense tantes limitacions.
Les tràquees dels artròpodes
En la majoria dels artròpodes (per exemple els insectes) la respiració té lloc a través de les tràquees, petits tubs ramificats que comuniquen amb l’exterior a través dels espiracles, orificis que solen obrir-se als costats del cos. Aquests canals minúsculs se subdivideixen fins a assolir un diàmetre microscòpic, i formen una xarxa que recorre tot el cos de l’animal i es va compactant a mesura que l’òrgan o el teixit corresponent són més actius. Normalment les ramificacions més fines són plenes de líquid.
L’oxigen i el diòxid de carboni circulen per les tràquees per difusió. Aquest sistema respiratori és una forma directa d’intercanvi gasós entre l’aire exterior i les cèl·lules i, per tant, és supèrflua la presència de sang com a vehicle dels gasos. Les parets de les tràquees són particularment primes, però al mateix temps disposen d’estructures específiques (petites plaques quitinoses en espiral, per exemple) que impedeixen que les parets s’enganxin i el canal quedi obstruït per la pressió d’altres òrgans (com, per exemple, els budells quan s’omplen). Sovint els espiracles presenten plomalls de pèls que impedeixen l’entrada de cossos estranys i gairebé sempre es poden tancar. De tota manera, les seves dimensions reduïdes són un sistema de defensa eficaç contra l’evaporació.
Les tràquees de les abelles i els diversos coleòpters presenten dilatacions que formen vesícules, les quals constitueixen reserves d’aire per al vol, moment en què la necessitat d’oxigen és més gran.
Els pulmons
L’altre sistema respiratori típic dels animals terrestres és el pulmonar. En aquest cas els intercanvis gasosos també tenen lloc en una cavitat interna del cos, el pulmó, per tal d’evitar l’evaporació. Tenen pulmons els gastròpodes, els amfibis, els rèptils, els ocells i els mamífers. Els pulmons dels animals menys complexos com els gastròpodes són simples cavitats del mantell revestides per una paret prima i amb abundants vasos sanguinis. Els dels amfibis i, sobretot, els rèptils són més evolucionats i la superfície d’intercanvi, notablement més pronunciada entre els rèptils, pot augmentar de maneres diverses com plegaments o, fins i tot, amb estructures esponjoses.
Els ocells presenten un augment extraordinari de la superfície pulmonar, cosa que permet una activitat respiratòria molt més intensa. Recordem que un ocell gasta durant el vol molta més energia que un mamífer que corre per terra. En aquests animals, l’aire circula per una llarga tràquea, reforçada per anells cartilaginosos, i després passa als bronquis, que es ramifiquen en els bronquíols. D’aquests bronquíols surten uns tubets petits de parets delicades (els bronquíols àcins) que constitueixen un reticle esponjós, a través del qual l’aire flueix com ho faria l’aigua per una esponja, que travessa completament. A banda dels veritables pulmons, els ocells tenen un complicat sistema de sacs aeris de parets primes que ocupen gran part de la cavitat toràcica i l’abdomen.
Aquests sacs també comuniquen amb els espais plens d’aire que hi ha a l’interior de gairebé tots els ossos dels ocells. Durant la inspiració la major part de l’aire omple els sacs aeris posteriors, després passa a través dels pulmons, on té lloc l’intercanvi respiratori, i finalment omple els sacs aeris anteriors, dels quals és expulsat a continuació. D’aquesta manera no solament s’assegura un flux continu d’aire a través dels pulmons, sinó que a més, en quedar-se als sacs anteriors l’aire per expirar, la taxa de diòxid de carboni es manté constantment alta, com també la freqüència respiratòria, que depèn directament de la concentració d’aquest gas.
Aquest conjunt pulmons-sacs aeris permet als ocells volar molt amunt. Mentre que els mamífers comencen a mostrar signes de manca d’oxigen entre els 3 000 i els 4 000 m, els ocells poden volar sense problemes a altituds superiors a 6 000 m (s’han donat casos d’avions que han topat amb ocells a 7 000 m). En efecte, el sistema respiratori dels ocells permet l’aprofitament òptim de l’oxigen present en l’aire a baixa pressió. Això és possible perquè els fluxos de l’aire i la sang a través dels pulmons segueixen direccions oposades. A mesura que l’aire travessa els pulmons i es dirigeix cap als sacs anteriors, cedeix cada cop més quantitat d’oxigen. La sang venosa, que avança en direcció contrària, troba aire amb concentracions d’oxigen cada cop més elevades. Es tracta d’un sistema molt semblant al de les brànquies dels peixos.
Els mamífers, com els ocells, tenen pulmons que comuniquen amb l’exterior a través de la tràquea i els bronquis, però en aquest cas els bronquíols acaben en unes estructures en forma de saquet, els alvèols, on tenen lloc els intercanvis gasosos. Els alvèols, amb un diàmetre aproximat d’1 mm, són coberts per una fina pel·lícula humida i envoltats per una xarxa compacta de capil·lars. Els pulmons no són exclusius dels organismes terrestres. Els peixos també presenten formes rudimentàries de pulmons. Molts peixos tenen sacs plens d’aire, les bufetes natatòries, connectades al sistema digestiu; l’aire és engolit i contingut en aquestes bufetes, a través de la paret de les quals tenen lloc els intercanvis gasosos.
Generalment, a més de les bufetes natatòries hi ha un sistema branquial, però els dos òrgans respiratoris poden ser emprats alternativament o fins i tot al mateix temps. Hi ha un tipus de peixos, els peixos pulmonats, que són considerats com la transició entre els peixos i els vertebrats terrestres, ja que estan mancats de brànquies i només tenen pulmons. Per tant, per a respirar han de sortir a la superfície periòdicament. Però, a diferència dels altres peixos, quan els torrents o els estanys on viuen s’assequen poden sobreviure fins a l’arribada de la propera estació de les pluges.
Un cas diferent és el dels amfibis. Durant les primeres etapes de la vida respiren a través de brànquies, que s’atrofien progressivament mentre la respiració esdevé preferentment, però no exclusivament, pulmonar. En efecte, la respiració cutània continua mantenint una importància molt rellevant.
La circulació en els diversos grups zoològics
Els animals unicel·lulars i els pluricel·lulars més simples com els porífers, els celenterats i els cucs són de petites dimensions i per tant els seus teixits i cadascuna de les cèl·lules del cos estan pràcticament en contacte directe amb les substàncies nutritives i amb l’oxigen procedents del medi. Els organismes més complexos i de dimensions més grans presenten, en canvi, un autèntic sistema de transport i distribució.
Aquest sistema consisteix en un conjunt de tubs per l’interior dels quals circula un fluid que funciona com a vehicle de tots els elements que han d’arribar als diversos teixits de l’organisme o que se n’han d’allunyar. Perquè la circulació del fluid per l’interior de l’organisme sigui contínua, un o més òrgans especialitzats tenen la missió de proporcionar-li rítmicament una empenta propulsiva.
Dins l’escala zoològica, els anèl·lids són els primers que presenten un sistema circulatori estructurat d’aquesta manera. És format per dos grans vasos (un de dorsal i un de ventral) que travessen tot el cos, per vasos circulars i capil·lars. La sang es manté en moviment gràcies a la contracció de les zones musculars del vas dorsal i dels vasos circulars, veritables cors. Es tracta d’un sistema circulatori tancat, en què la sang sempre roman a l’interior dels vasos. L’intercanvi de substàncies nutritives, de rebuig i d’aigua té lloc a través de la paret dels capil·lars.
En els mol·luscs i els artròpodes, en canvi, la circulació és oberta. La sang pot fluir a través de l’exterior dels vasos directament als teixits. El cor dels mol·luscs és petit i situat dorsalment. En general té dues cavitats, els atris, que reben la sang procedent de les brànquies o dels pulmons i l’empenyen cap a les artèries. Des d’allà la sang passa a les llacunes de la cavitat general del cos i successivament es recull en les venes que la porten als òrgans respiratoris, on, abans de tornar al cor, absorbeix més oxigen. Els cefalòpodes també tenen cors suplementaris, situats a la base de les brànquies.
Els insectes tenen un sistema circulatori més aviat reduït. En aquests animals l’oxigen no necessita la sang com a vehicle, perquè s’escampa pels teixits del cos a través de les tràquees. Així, el cor, en forma de tub i en posició dorsal, bomba la sang cap a la cavitat del cos mitjançant una sola aorta curta, orientada cap endavant. No hi ha artèries laterals ni capil·lars ni venes, i la sang (que no conté cap pigment respiratori i s’anomena hemolimfa), després d’irrigar els diversos òrgans i teixits, es dirigeix al cor gràcies al batec de dues membranes de l’abdomen i, després, és aspirada de nou pel cor quan aquest és dilatat pels músculs de les ales.
El sistema circulatori de tots els vertebrats és tancat. Com en el cas dels anèl·lids, la sang sempre roman a l’interior dels vasos i tots els intercanvis cel·lulars de materials i de gasos tenen lloc a través de la paret dels capil·lars. Dins el mateix grup s’observen diverses modificacions sensibles de l’aparell circulatori, estretament relacionades amb els processos d’adaptació al medi.
Sobretot en els vertebrats terrestres, el cor es modifica i canvia la mateixa estructura del sistema, que fins i tot es desdobla en dos circuits diferents: l’un s’encarrega de distribuir la sang amb oxigen abundant a totes les parts de l’organisme i l’altre té la missió de conduir-la cap als òrgans respiratoris.
El sistema circulatori dels peixos encara s’estructura en un únic circuit: el cor, que només és format per dues cavitats (un atri i un ventricle). Rep només sang venosa i la bomba directament a les brànquies, a través d’un gran vas del qual surten 4 parells d’artèries branquials i nombrosos capil·lars. Les grans artèries distribueixen la sang oxigenada a totes les parts del cos.
En els amfibis ja se’n diferencien dos circuits: el pulmonar i el general, però això no vol dir un increment de l’eficàcia del sistema. El cor té dos atris, l’un és recorregut per sang venosa procedent del circuit general i l’altre per sang molt oxigenada procedent dels pulmons. Però només hi ha un sol ventricle, per la qual cosa les sangs es barregen inevitablement. El resultat és que en el circuit general es bomba sang que, en conjunt, té una taxa d’oxigen més aviat baixa. Tanmateix, com que bona part dels intercanvis respiratoris es realitzen a través de la pell, aquest desavantatge no resulta gaire rellevant per als amfibis.
La situació és ben diferent per als rèptils, que, adaptats a la vida terrestre i amb l’epidermis consegüentment engruixida, han perdut la capacitat de respirar per via cutània. Per a millorar l’eficàcia del sistema, el seu cor ha hagut de modificar-se forçosament per tal de mantenir separades les dues sangs. Així, a l’interior del ventricle s’ha creat una paret divisòria, encara incompleta, gràcies a la qual s’aconsegueix que la sang que circula pel circuit general sigui preferentment l’oxigenada que prové del circuit pulmonar.
Solament els animals amb temperatura interna constant, és a dir, els ocells i els mamífers, presenten separació total dels dos ventricles i per tant separació de la circulació pulmonar i la general. Des del ventricle dret s’empeny cap als pulmons la sang pobra d’oxigen, mentre que del ventricle esquerre i en direcció a tots els teixits de l’organisme en surt exclusivament sang oxigenada.
La nutrició
Tots els éssers vius, unicel·lulars i pluricel·lulars, vegetals i animals, tenen algunes funcions essencials que són en conjunt les que els proporcionen la vida i els distingeixen dels éssers no vius. Una d’aquestes funcions és la nutrició, que en els animals se satisfà de maneres molt diverses, de vegades fins i tot curioses. Els únics òrgans dels quals estan mancats són els necessaris per a la fotosíntesi.
Les estructures bucals
L’alimentació dels animals és extremament variada: des de llana fins a terra, passant per carronya, pol·len, herba, mel, fusta i sang. El primer requisit és tenir l’"instrumental" adequat per a introduir els aliments a l’interior del cos. La boca pot adoptar les estructures més diverses i de vegades estranyes. Per exemple, els animals que es nodreixen d’aliments líquids poden tenir petites probòscides, normalment enrotllades en espiral, que després estiren per xuclar (tal com fan les papallones que s’alimenten de nèctar), o bé estructures en forma d’agulles (és el cas dels mosquits), o bé una obertura en forma de ventosa proveïda de dents còrnies per a aferrar-se a la presa i xuclar-li la sang (és el cas de la llamprea).
Hi ha casos en què les dents són molt importants en el procés de la ingestió de l’aliment. Les dents dels mamífers carnívors, per exemple, són esmolades i punxegudes i no serveixen tant per a mastegar com per a retenir, amb l’ajut de les urpes, la presa i trossejar-ne la carn. Aquesta mateixa funció, lacerar la carn, és realitzada entre els ocells rapinyaires per un bec corni i robust.
Els animals que es nodreixen de plantes, en canvi, han de resoldre el problema de la resistència especial que presenten els teixits vegetals a causa de l’existència de cel·lulosa. Els mamífers resolen aquest problema amb dents planes i proveïdes de petits solcs que funcionen com veritables moles.
La llengua també pot adoptar formes i funcions diverses. Alguns mamífers que es nodreixen de formigues tenen la llengua llarga, prima i viscosa, així es pot ficar per l’interior del formiguer i capturar centenars de formigues de cop. En altres casos, com la granota i el camaleó, és un veritable estri per a caçar. Estimulada per les preses en moviment, la llengua surt disparada de la boca i atrapa l’insecte a l’instant, a tal velocitat que no és perceptible ni per l’ull humà ni pel de la majoria dels animals.
El processament dels aliments
Un cop l’aliment ha estat incorporat, ha de ser sotmès a una sèrie de tractaments, tant mitjançant la fragmentació mecànica que realitzen els músculs de la paret de l’aparell digestiu com per l’acció química, fins que es redueix a molècules simples que poden travessar les membranes cel·lulars per a ser utilitzades per les cèl·lules.
Els animals més simples, els protozous unicel·lulars, no tenen ni boca ni aparell digestiu ben diferenciats. L’ameba, per exemple, engloba l’aliment (fragments de material orgànic vegetal o animal, altres protozous) amb pseudòpodes, prolongacions mòbils del citoplasma, i l’introdueix al cos, on forma un vacúol digestiu. A l’interior del vacúol digestiu intervenen enzims que digereixen les substàncies nutritives, les quals després, reduïdes a dimensions mínimes, poden passar al citoplasma a través de la membrana del vacúol. Els protozous que s’alimenten de partícules orgàniques més grosses han desenvolupat una forma de digestió extracel·lular, que és característica dels animals més evolucionats: els enzims són alliberats fora de la cèl·lula i predigereixen l’aliment abans d’absorbir-lo. Els protozous ciliats, anomenats així perquè a la superfície cel·lular tenen estructures mòbils que recorden pèls i que els permeten el moviment, ja presenten diferenciació rudimentària de la boca i l’anus. Tenen una zona especial a la paret cel·lular que s’estova quan l’aliment passa al citoplasma, on es forma el vacúol digestiu. A més, hi ha una altra zona de la membrana cel·lular que fa d’anus.
En els celenterats, que inclouen les meduses, les actínies o anemones de mar, etc., la boca i l’anus coincideixen, és a dir que fan servir la mateixa obertura per a introduir l’aliment i per a eliminar els residus no digerits. Aquests animals tenen el cos en forma de sac, el qual comunica amb l’exterior per mitjà d’una obertura envoltada de tentacles mòbils que serveixen per a capturar la presa, a la qual inoculen un verí, i ficar-se-la a la "boca". A l’interior de la cavitat del sac, o cavitat gastrovascular, té lloc la digestió, en part extracel·lular (efectuada mitjançant els enzims alliberats per cèl·lules particulars de la paret) i en part intracel·lular, algunes cèl·lules es desprenen de la paret i engloben i digereixen, com els protozous, les partícules d’aliment. Novament en contacte amb els teixits de la cavitat gastrovascular, sembla que cedeixen a les cèl·lules del voltant alguns productes de la digestió.
Com els celenterats, els cucs plans (platihelmints) tenen una única obertura que comunica amb l’exterior i una cavitat digestiva indiferenciada que s’estén, amb ramificacions, a quasi tot el cos de l’animal. Els cucs cilíndrics (nematohelmints), en canvi, són els primers que presenten un veritable aparell digestiu, per bé que rudimentari. Consisteix en un tub que recorre tot el cos de l’animal, i als extrems del qual es pot distingir una boca (en alguns casos proveïda de dents capaces d’agafar l’aliment) i un anus.
Entre els anèl·lids, grup de cucs que difereix dels cilíndrics perquè el cos és segmentat, és a dir, format per una sèrie d’unitats cilíndriques unides les unes a les altres, el sistema digestiu és constituït per un tub que recorre el cos en tota la seva llargada, de la boca a l’anus, però que se subdivideix en parts especialitzades on es produeixen els diversos processos digestius. En el cuc de terra, per exemple, es reconeix un veritable estómac, dilatació del tub digestiu, on roman l’aliment temporalment i on és barrejat i triturat pels moviments musculars de la paret. En aquest nivell ja s’observen les dues característiques fonamentals dels sistemes digestius dels animals més evolucionats: la digestió extracel·lular, que té lloc en una cavitat del cos preparada per a aquesta finalitat específica, i l’especialització d’aquesta digestió en diversos òrgans com l’estómac i les diferents parts de l’intestí, cadascuna de les quals serveix per a una tasca concreta.
La divisió de les tasques en l'aparell digestiu
La divisió del treball en l’aparell digestiu garanteix una eficàcia molt més gran a tot el sistema, però no tots els òrgans fan les mateixes funcions en els diferents animals. En general, la digestió comença amb la masticació, que és una primera fragmentació de l’aliment, però en els ocells això no és possible ja que no tenen dents òssies. Aquesta funció és assumida per l’estómac, a l’interior d’una saca muscular especialitzada anomenada pedrer, que amassa i tritura l’aliment. Un cas curiós és el de l’estómac dels ocells granívors, com per exemple les gallines, que sovint conté pedretes que, en barrejar-se amb l’aliment, potencien l’acció trituradora de les parets musculars de l’estómac.
Un altre exemple d’estómac especial és el dels herbívors que es nodreixen de vegetals particularment durs i resistents, com les tiges del blat i el blat de moro. Tenen una digestió llarga, perquè cal molt de temps perquè la cel·lulosa (component principal dels organismes vegetals) sigui triturada del tot. Per tant, l’estómac dels herbívors és particularment desenvolupat i dividit. Una part de l’estómac és ocupada immediatament per l’aliment, pràcticament intacte, ingerit durant la pastura. Després l’aliment serà sotmès a un llarg procés de masticació, un cop regurgitat de l’estómac a la boca (ruminació).
En una altra part de l’estómac té lloc la veritable digestió química. En general, la velocitat d’aquesta digestió depèn del grau de fragmentació de l’aliment, perquè les partícules nutritives tan sols poden ser atacades per la superfície. Com més petits són els fragments d’aliment, més gran és la superfície atacable pels enzims digestius i, per tant, la velocitat del procés digestiu és superior. El conjunt d’enzims presents a l’estómac comença l’escissió de les proteïnes. Aleshores l’aliment passa a l’intestí prim a través d’una vàlvula muscular que no falta gairebé mai, anomenada esfínter pilòric i situada al fons de l’estómac.
L’intestí és la part de l’aparell digestiu on es produeix l’absorció de les substàncies nutritives, és a dir, on passen a la sang, un cop acabada la digestió química. En general és ple de vasos sanguinis que poden transportar a les diverses parts del cos les molècules absorbides. El fragment intestinal anomenat intestí gros és ple de microorganismes i en molts animals hi ha uns sacs anomenats "cecs" que es ramifiquen des del fragment principal, on romanen durant cert temps les restes alimentàries i on són atacades pels bacteris. Els cecs són més desenvolupats en els animals herbívors, mentre que els carnívors tenen una digestió menys laboriosa i tubs digestius més curts i lineals. Les matèries fecals s’acumulen a la part final de l’intestí, el recte, des d’on són eliminades periòdicament.
La digestió de la cel·lulosa
Dos animals tan diferents com una vaca (mamífer) i un tèrmit (insecte) tenen una característica fisiològica comuna: tots dos basen la seva alimentació en la digestió de la cel·lulosa, component principal del farratge i de la fusta. El seu estómac funciona com un fermentador industrial molt eficient. És un microcosmos anaeròbic amb una comunitat microbiana molt activa, que transforma la cel·lulosa en compostos químics assimilables per l’animal que l’aixopluga. La cel·lulosa és un sucre de pes molecular elevat format per cadenes de molècules de glucosa unides mitjançant enllaços químics. Així com la glucosa és assimilable pels animals, molt pocs organismes disposen de les eines necessàries —els enzims— per a trencar els enllaços químics que uneixen les molècules de cel·lulosa. Aquests organismes comprenen algunes espècies de fongs, protoctists i, principalment, bacteris.
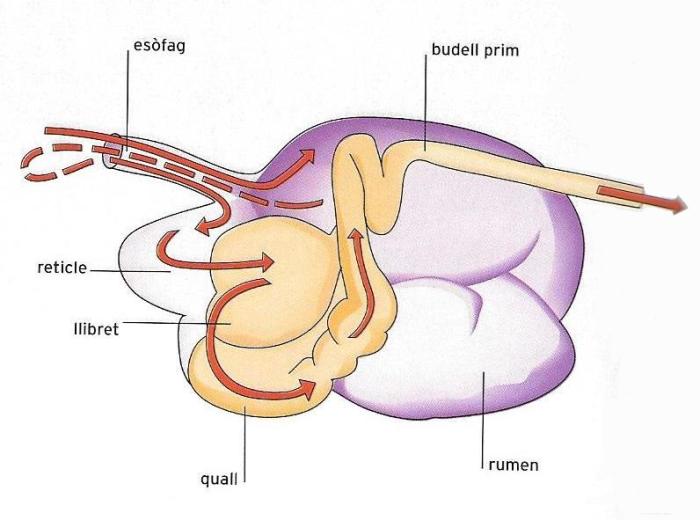
Esquema del rumen i el sistema gastrointestinal de la vaca. El menjar prové de l'esòfag i va a parar al rumen; després és regurgitat i passa pel llibret, el quall i els budells, per aquest ordre.
ECSA
L’aparell digestiu d’un degradador de cel·lulosa és una mena de ciutat microbiana els habitants de la qual han evolucionat conjuntament entre ells i amb l’animal que els acull, establint-hi una relació simbiòtica en què cadascun dels participants acompleix una tasca específica. En el cas dels remugadors, l’animal prepara l’aliment mastegant-lo i barrejant-lo amb abundant saliva (una vaca pot produir-ne entre 60 i 100 litres per dia) fins a formar el bol alimentari, que és engolit i passa a la primera cavitat de l’estómac, anomenada rumen (vegeu el dibuix). Allà, els fragments vegetals són atacats pels microbis; de manera activa, en el cas de microorganismes mòbils, o per adherència a la superfície dels teixits, en el cas dels bacteris sense motilitat. L’animal contreu les parets del rumen, cosa que afavoreix la mescla del contingut i, per tant, la digestió. Després regurgita l’aliment mig paït i el mastega per segona vegada (remugament), fragmentant-lo encara més i untant-lo amb més saliva. Les partícules més petites i denses del bol alimentari, en retornar a l’estómac passen a una altra cavitat (reticle).
En el rumen hi ha bacteris que trenquen les cadenes de cel·lulosa i n’obtenen sucres més senzills, que són fermentats per altres microbis. Alguns dels productes de la fermentació serveixen encara de font d’energia a altres microorganismes. Entre els productes finals destaquen els àcids grassos, que són absorbits per l’epiteli de l’estómac i s’incorporen al metabolisme de l’animal. També es produeix una quantitat significativa de metà (CH4) i de diòxid de carboni (CO2), gasos que surten a l’exterior mitjançant eructes. A més de bacteris, que en són els principals components, la microbiota del rumen conté protoctists —gairebé exclusivament ciliats— i fongs, que solen alternar la forma flagel·lada i la de tal·lus. Els fongs poden metabolitzar parcialment altres polisacàrids, a més de la cel·lulosa, com ara la lignina, l’hemicel·lulosa i les pectines. La pròpia microbiota serveix d’aliment al remugant; els seus components són digerits en la darrera cavitat de l’estómac, que és àcida, i són una font de proteïna per a l’animal. Els cavalls i els conills són herbívors, però no tenen rumen. El seu estómac és senzill, i la microbiota cel·lulòsica, que fa la seva tasca en una cavitat del budell gros, molt a prop de l’anus, és expulsada amb els excrements. No s’aprofita, per tant, com a font de proteïnes. Els conills solen recuperar-ne una part menjant els propis excrements.
L’estómac de les balenes també té diversos compartiments, i hi té lloc una fermentació semblant a la del rumen. Un dels components bàsics de l’alimentació d’aquests mamífers marins és el krill, conjunt de petits crustacis planctònics que, com tots els artròpodes, tenen una coberta de quitina, la qual és metabolitzada per la microbiota simbiòtica de la balena.
Entre els ocells, n’hi ha un que té el pap adaptat per a la fermentació cel·lulòsica. És l’hoatzin, un ocell tropical que s’alimenta exclusivament de fulles; la cel·lulosa és, per tant, la seva única font de carboni.
Hi ha també insectes que contenen microorganismes simbiòtics que degraden la cel·lulosa, com ara diverses paneroles xilòfagues (menjadores de fusta), les larves d’alguns coleòpters, o els tèrmits (isòpters), que amb uns dos milers d’espècies pertanyents a 210 gèneres constitueixen el principal grup d’insectes especialitzat a menjar fusta. Viuen a les sabanes, als boscos tropicals, a les praderies i als deserts, i s’alimenten exclusivament de cel·lulosa, excepte els individus del gènere Mastotermens, que, a més de fusta, mengen qualsevol altra cosa que contingui carboni, fins i tot pneumàtics o el recobriment de cables elèctrics. Els tèrmits tenen una panxa voluminosa on viuen comunitats microbianes d’una gran complexitat. Inclouen centenars d’espècies diferents de protoctists, alguns dels quals tenen també bacteris simbiòtics en el seu aparell digestiu, i cada espècie de tèrmit té els seus protoctists específics. Els bacteris també són molt abundants a l’aparell digestiu dels tèrmits xilòfags, però no se n’ha aïllat cap que degradi la cel·lulosa.
Les glàndules digestives
Les glàndules digestives són molt importants en tots els sistemes digestius. Poden ser simples cèl·lules que entapissen la cavitat digestiva i hi introdueixen directament els enzims, o bé es poden organitzar com veritables òrgans (com les glàndules salivals, el fetge i el pàncrees dels vertebrats) que aboquen els seus productes al tub digestiu. En parlar de la secreció de les glàndules, cal remarcar que el mecanisme estímul/resposta és fonamental per al funcionament del sistema digestiu de tots els animals en general. En efecte, és l’olor i l’aspecte del menjar allò que n’estimula la captura i la ingestió, i és la presència d’aliment en el tub digestiu allò que estimula els moviments musculars i la secreció enzimàtica.
L'excreció
En les cèl·lules animals tenen lloc ininterrompudament processos de trituració i d’elaboració de substàncies químiques, amb una producció contínua de residus. Aquests residus són eliminats constantment, perquè si s’acumulessin en l’organisme seria molt nociu.
Entre els residus que cal eliminar hi ha el diòxid de carboni (CO2), que és absorbit principalment pels pulmons; els productes terminals de la degradació de les proteïnes, compostos de nitrogen com la urea i l’àcid úric, eliminats a través de l’orina; i també hi ha excés d’aigua, que surt a l’exterior a través de l’aparell respiratori, l’intestí, la pell (en els ungulats i els simis en forma de suor) i, sobretot, a través de l’orina, líquid de composició variable elaborat per determinats òrgans excretors.
La necessitat d'un equilibri intern estable
La funció fonamental del sistema excretor és regular i mantenir constant l’homeòstasi, és a dir, les condicions d’equilibri de l’interior de l’organisme. L’eliminació dels residus és només un dels aspectes importants d’aquest sistema. Els diversos mecanismes biològics cel·lulars, per a poder mantenir la seva eficàcia, han de treballar en un medi on les característiques siguin constants; on, per exemple, les concentracions dels diversos elements i compostos químics siguin substancialment estables. Tots els animals, doncs, llevat dels organismes marins més simples en els quals l’ambient intern és pràcticament igual que l’extern (els seus fluids orgànics són molt semblants a l’aigua de mar), han d’exercitar una regulació rigorosa de les condicions internes que mantingui constants les característiques. Per exemple, la concentració de sals dels fluids interns d’un animal que viu en aigua dolça és normalment molt superior a l’exterior. En una situació com aquesta, si no intervinguessin altres factors, pel fenomen de l’osmosi tindria lloc la difusió d’aigua a l’interior del cos de l’animal, de manera que es produiria un inflament progressiu que conduiria a la mort. Els òrgans excretors eviten aquest perill eliminant constantment l’excés d’aigua. Aquesta eliminació té lloc bé per l’expulsió a l’exterior d’una orina molt diluïda, elaborada en òrgans especialitzats, o bé, en els casos més simples com els protozous d’aigua dolça o les esponges, per la intervenció d’òrgans excretors, rudimentaris però eficaços, com els vacúols contràctils. Aquests s’omplen d’aigua que inclou substàncies de rebuig dissoltes i es buiden periòdicament abocant-la a l’exterior.
Els òrgans excretors dels cucs plans, anomenats protonefridis, estan formats per dos canals longitudinals que desemboquen directament a l’exterior; reben nombrosos túbuls laterals de fons cec, en els quals es filtra el líquid d’excreció, que plomalls de cilis fan avançar contínuament. En els anèl·lids, en canvi, hi ha un parell de túbuls excretors per a cada segment, els metanefridis, que s’obren al celoma mitjançant un embut ciliat, a través del qual passen les substàncies de rebuig. El més curiós d’aquest sistema és la presència de cèl·lules migratòries que transporten els residus i després es desintegren en el celoma.
Els insectes tenen una sèrie de túbuls llargs i prims que es ramifiquen per tot el cos i s’obren a la part posterior de l’intestí. Les substàncies de rebuig passen a través dels túbuls de la sang a l’aparell digestiu i des d’aquí cap a l’exterior. Abans que l’orina sigui expulsada es reabsorbeixen algunes substàncies útils per a l’organisme. En moltes espècies, l’orina és recollida en una bufeta que es buida de tant en tant.
Els ronyons dels vertebrats
Els òrgans principals de l’aparell excretor dels vertebrats són els ronyons. Encara que varien d’una espècie a l’altra, els ronyons, òrgans parells, sempre presenten la mateixa estructura bàsica: un amàs de túbuls que desemboquen en un mateix conducte col·lector. La majoria dels túbuls renals s’origina en una petita càpsula (anomenada de Bowman) on s’apleguen un garbuix de vasos sanguinis. Els túbuls es repleguen i s’entrellacen i després convergeixen en un conducte, l’urèter, que va a parar a la bufeta, connectada amb l’exterior per mitjà d’un conducte anomenat uretra. En tots els túbuls excretors, des dels més rudimentaris dels cucs fins als dels mamífers, el procés de formació de l’orina és bàsicament el mateix: al començament del túbul, el líquid contingut és molt semblant al sèrum de la sang o als fluids interns. Però mentre passa pel túbul aquesta solució es va processant, s’hi afegeixen algunes substàncies i se’n recuperen d’altres, i al final, el líquid que s’elimina a l’exterior, l’orina, és diferent dels fluids interns.
Els humans poden sobreviure molt poc temps sense aigua dolça. Eliminen grans quantitats d’aigua a través tant de la sudació com de l’orina, i en condicions de manca d’aigua entren ràpidament en estat de deshidratació mortal. En canvi, hi ha altres mamífers que poden viure amb quantitats d’aigua dolça decididament exigües. Són animals adaptats a la vida en medis àrids i els seus ronyons tenen gran capacitat de concentració, és a dir, produeixen una orina molt poc diluïda i d’aquesta manera limiten la pèrdua d’aigua.
La rata del desert, per exemple, estalvia l’aigua produint petites quantitats d’orina extremament concentrada: la concentració de sodi és sis vegades superior a la del plasma, i la concentració d’urea és quatre vegades més gran que la de l’orina humana. Així pot sobreviure fins i tot sense beure, només amb l’aigua que contenen els aliments sòlids i la que resulta com a subproducte en els processos metabòlics.
Estructures de revestiment i sosteniment
En tots els animals, la capa més externa és el límit entre l’individu i el medi. En els primers organismes vius aquesta capa, l’epidermis, era el centre d’intercanvi dels gasos, on es difonien els materials de rebuig, es registraven les variacions externes i era possible el moviment (a través dels cilis).
Els diversos tipus d'epidermis
En els animals poc evolucionats l’epidermis manté algunes de les funcions originàries que hem citat. Per exemple en el cuc de terra, les larves dels crustacis branquiòpodes i en altres grups la pell encara funciona com a òrgan respiratori. En els animals amb organització més complexa, en canvi, moltes funcions han estat transferides a teixits i òrgans especialitzats, i la funció més important de la pell és la de constituir una barrera protectora entre l’interior i l’exterior.
Al llarg del temps l’epidermis s’ha hagut d’adaptar a medis molt diversos i ha desenvolupat formacions determinades com els pèls, les plomes, les escates dels vertebrats i les rígides estructures de protecció dels invertebrats. Així, per exemple, l’epidermis dels coralls produeix un esquelet calcari que té funció de protecció i de suport alhora, mentre que les cèl·lules cutànies d’alguns cucs secreten un moc que aglutina els granets de sorra i els trossets de petxines per construir-se al voltant del cos una mena d’escut rígid de protecció.
Els mol·luscs tenen cèl·lules especials a l’extrem del mantell que "construeixen" un sistema de suport i protecció molt particular, la closca. Les closques tenen formes i estructures molt variades. Poden ser compostes d’una sola peça o de dues, empernades entre si, afuades o planes, amb espirals sobre el mateix pla o enrotllades sobre elles mateixes, amb les primeres voltes de l’espiral tancades del tot per les últimes. En alguns gastròpodes la closca és molt petita i recoberta del tot pel mantell; en molts casos, com el dels nudibranquis, és del tot absent. Les closques dels mol·luscs, com el tub protector dels anèl·lids, són secretades només per una part de l’epidermis, i per això es distingeixen de la cutícula, el revestiment protector produït per totes les cèl·lules cutànies. La cutícula més evolucionada és la dels artròpodes, i és un revestiment ric en quitina. La cutícula dels crustacis és calcària. Hi ha zones endurides alternades amb zones flexibles: el resultat és un exosquelet articulat i elàstic que protegeix els òrgans interns, aguanta el cos i, en proporcionar un lloc de fixació dels músculs, permet moviments que poden arribar a ser complexos. L’exosquelet dels artròpodes, que embolcalla tot el cos, és una estructura molt perfeccionada, però té una greu limitació; un cop format és estable i, per tant, la seva mida no pot continuar augmentant a mesura que l’animal creix. En un determinat moment del creixement, l’exosquelet queda massa petit i l’animal se n’ha de desprendre per a fer-se’n un de més gran. És el procés de la muda, que té lloc periòdicament.
L'epidermis dels vertebrats
La pell dels vertebrats és més aviat gruixuda i composta per diverses capes de cèl·lules. En molts casos s’ha diferenciat per a donar lloc a formacions molt particulars. En els peixos ossis, per exemple, l’epidermis ha originat les escates, primes plaques flexibles, riques de sals de calci. Encara que no és fàcil de veure, les escates queden cobertes per la cutícula, que s’adapta periòdicament al creixement de l’animal.
L’epidermis dels rèptils és molt gruixuda i conté una substància proteica, la queratina, que a més de protegir l’organisme evita que es deshidrati. No s’ha d’oblidar que van ser precisament els rèptils els primers animals que s’adaptaren al medi terrestre. Els cocodrils i les tortugues es protegeixen amb plaques còrnies, mentre que les sargantanes i les serps tenen escates més petites, imbricades i, al cap, escuts cornis. Com que tenen aquest revestiment tan poc extensible, també els rèptils per a poder créixer han de patir periòdicament el procés de la muda.
Els ocells posseeixen escuts cornis semblants a les potes i, modificats, al bec. Les mateixes plomes són una estructura derivada de les escates dels rèptils. Les funcions de les plomes dels ocells són nombroses: formen un revestiment resistent i impermeable a l’aigua i, en retenir l’aire, creen una mena de coixinet aïllant al voltant de l’animal. Els ocells que viuen a l’aigua tenen una glàndula particularment desenvolupada, l’uropigi, que secreta un oli que impermeabilitza les plomes. Un o dos cops l’any, segons un cicle hormonal, les plomes són substituïdes, en part o del tot, per altres plomes noves. En moltes espècies de les zones temperades la muda té lloc dos cops l’any i així es diferencia el vestit estival de l’hivernal. En nombrosos casos, el mascle incorpora durant l’època reproductiva una vestimenta nupcial, amb un plomatge sovint abundant i vistós.
La formació epidèrmica més característica dels mamífers són els pèls, que protegeixen l’animal, li donen color i conserven l’escalfor del cos. Molts mamífers, com el lèmming o la llebre, pateixen una mena de muda dos cops l’any, a la primavera i la tardor, i d’aquesta manera l’animal pot disposar d’un pelatge en alguns casos críptic (blanc a l’hivern) de pèls atapeïts i llargs durant els mesos més freds, i d’un pèl més curt durant els mesos calorosos. Sembla que el fenomen extern que determina el començament de la muda és la variació de la durada de la llum diürna.
Les estructures de sosteniment
Tots els animals necessiten sistemes de sosteniment per a evitar que el cos caigui sota l’acció de la força de gravetat. Són una excepció alguns organismes aquàtics com les meduses o les actínies, ja que la seva consistència els permet conservar la forma a l’aigua sense cap estructura especial.
En els invertebrats és l’epidermis que assumeix, en molts casos, la funció de protecció i suport del cos, i de vegades, tal com hem vist, amb formacions molt característiques. Els vertebrats, en canvi, es caracteritzen perquè tenen un sistema de sosteniment intern, un endosquelet que també serveix de suport rígid d’inserció dels músculs. Conté dos teixits especialitzats: l’os, ric en sals minerals i en calci, i el cartílag, flexible i elàstic. Entre els adults, el cartílag tan sols apareix als extrems dels ossos, i entre els joves forma tot l’esquelet, però el teixit ossi el va substituint progressivament. Però hi ha alguns peixos, com els taurons i les rajades, que tenen l’esquelet format només de cartílag fins i tot quan són adults, i per això s’anomenen peixos "cartilaginosos".
L’esquelet de tots els vertebrats presenta més o menys la mateixa estructura: el crani, l’involucre rígid on s’allotja el cervell; la columna vertebral, que protegeix la medul·la espinal; la caixa toràcica, formada per les costelles, i, en la majoria dels casos, dos parells d’extremitats que de vegades poden ser modificats, com les ales dels ocells i les aletes dels peixos.
Alguns vertebrats tenen, a més de l’endosquelet, un esquelet extern, el dermatosquelet. És el cas de les tortugues, revestides per un involucre molt robust, i dels cocodrils, coberts de plaques òssies.
El moviment
Com ja s’ha dit més amunt, una de les funcions principals de l’exosquelet és proporcionar una estructura rígida perquè els músculs s’hi insereixin, sobretot els destinats al moviment. L’esquelet, en anar-se adaptant al llarg del temps als medis i sistemes de locomoció diferents, ha patit modificacions i adaptacions que l’han fet cada vegada més especialitzat. Els peixos, per exemple, que han de moure’s en un medi 800 vegades més dens que l’aire, tenen un esquelet flexible, que pot afavorir els moviments sinuosos de la natació, i alhora sòlid i robust, per a poder suportar sense deformar-se empentes musculars excepcionalment potents.
Paral·lelament a l’especialització de l’esquelet, per a cada tipus de locomoció s’han desenvolupat característiques específiques. Els peixos, per exemple, presenten una sèrie de característiques que faciliten el moviment a l’aigua: una silueta hidrodinàmica, que permet minimitzar la resistència que ofereix el medi aquàtic, una pell impregnada de mucositat que redueix el fregament, una musculatura capaç de proporcionar al cos un vigorós impuls propulsor, i una sèrie d’aletes, amb infinites variants en les diferents espècies, cadascuna de les quals està destinada a una funció exacta relacionada amb l’equilibri, l’estabilitat i la propulsió.
Per nedar, la majoria dels peixos contreuen rítmicament els músculs ventrals, cosa que provoca ondulacions del cos i la cua que proporcionen un potent impuls en l’aigua. L’aleta dorsal i l’anal actuen generalment d’estabilitzadors; l’aleta caudal fa d’òrgan propulsor; les aletes ventrals contribueixen a mantenir el cos en equilibri, mentre que les pectorals tenen funcions diverses: serveixen d’instruments de precisió i determinen els moviments lents, com si actuessin de timó de profunditat.
Uns altres animals amb una locomoció i una estructura física del tot particulars són les serps. Com que no tenen potes, tot el cos participa del moviment. Hi ha un desenvolupament extraordinari de la massa muscular, composta de milers de músculs, i un desenvolupament longitudinal de l’animal igualment extraordinari, amb una columna vertebral que té fins a 400 vèrtebres i és molt flexible, gràcies a una característica única: la capacitat de cada vèrtebra de girar 50° sobre ella mateixa.
Les serps presenten diferents tipus de locomoció. Les serps llargues i primes es mouen amb una ondulació lateral: els músculs, contrets, exerceixen una força lateral sobre els possibles punts de suport del terreny (les pedres, els rocs, els matolls). A la pràctica, el rèptil s’hi recolza per allunyar-se’n. Les ones de contracció es propaguen per tot el cos i la serp adopta el "moviment de reptació" més sinuós possible per aprofitar el major nombre de punts de suport on fer pressió. Les serps grosses, en canvi, realitzen un desplaçament rectilini. Exerceixen forces adreçades cap avall i, per tant, han d’arrossegar-se, per a la qual cosa empren grups d’escates cutànies alternades.
També l’esquelet dels ocells ha patit enormes modificacions, en aquest cas per a adaptar-se cada vegada millor a les exigències d’un sistema de locomoció tan especial com el vol. Els ossos s’han buidat per a omplir-se d’aire i d’aquesta manera fer l’esquelet especialment lleuger (el de la fragata, ocell amb una envergadura de més de 2 m, pesa uns 110 g). Les mandíbules i les dents, massa pesants, han estat substituïdes pel bec, sòlid i molt lleuger. La massa muscular s’ha fet més densa, sobretot a la part central del cos on una cinquantena de músculs estan destinats al moviment de les ales. Una massa muscular tan poderosa requereix una base d’"ancoratge" excepcionalment resistent: la carena, una cresta òssia robusta que surt de l’estern. La caixa toràcica i la columna vertebral, en canvi, han perdut la flexibilitat. Les ales, un instrument de vol molt perfeccionat, són articulades, resistents i flexibles alhora. En general l’ala d’un ocell té la superfície superior convexa i la inferior còncava: d’aquesta manera es crea per sobre una succió d’aire que dóna impuls cap amunt, i a la cara inferior es crea una força que també va dirigida cap amunt i que contribueix a l’enlairament. Les ales s’articulen per tal de poder executar moviments de descens i d’enlairament, i poden canviar el perfil i la superfície reunint o desplegant les plomes.
Es podria concloure que, en comparació, els vertebrats terrestres de quatre potes no tenen problemes. En realitat no és així, ja que coordinar el moviment de quatre potes per tal d’aconseguir una locomoció alhora estable i ràpida, i que comporti la mínima despesa d’energia, no és tan senzill. Les potes han significat la gran adquisició evolutiva dels artròpodes i els vertebrats tetràpodes (és a dir, proveïts de quatre potes), i els han permès, mitjançant la deambulació, conquerir i colonitzar les terres emergides. Les potes dels artròpodes han evolucionat en relació amb un esquelet extern a les masses musculars i inicialment van aparèixer com a òrgans natatoris. En els vertebrats, en canvi, les potes han evolucionat en relació amb un esquelet intern a les masses musculars i com a resposta a la necessitat de moure’s per les terres emergides. És gràcies a l’adquisició de les potes, òrgans susceptibles d’originar adaptacions molt variades i complexes en funció dels usos més diversos (per a la cursa, per al salt, per a excavar, per a atrapar la presa, etc.), que els artròpodes i els vertebrats tetràpodes han sofert una evolució refinada i complexa de la musculatura i el sistema nerviós gràcies a la qual han pogut imposar-se a tots els altres grups animals. La locomoció terrestre es basa en les lleis de la palanca: les extremitats es mouen endavant, recolzen a terra i després impulsen el cos endavant gràcies a la resistència del sòl. El moviment de propulsió és seguit d’una fase de retorn, en què les extremitats tornen a recolzar sobre el terra.
El tipus de locomoció més estesa entre els mamífers terrestres és l’anomenada "en diagonal": l’extremitat anterior esquerre i la posterior dreta o l’extremitat anterior dreta i la posterior esquerra s’alcen i es desplacen endavant simultàniament. En definitiva, només hi ha dues potes recolzades al terra al mateix temps i el cos es manté dret per inèrcia, en una mena d’equilibri dinàmic.
En canvi, els animals amb les extremitats llargues, com la girafa i el camell, tenen una manera diferent de caminar, perquè aixequen alhora les dues potes del mateix costat, mentre les altres dues es mantenen recolzades. Això és el que dóna a aquests animals el característic caminar oscil·lant.