La cèl·lula eucariota: forma i estructura
Una ceba és constituïda d’esquames carnoses i concèntriques anomenades bràctees, que contenen aigua i substàncies de reserva en abundància. Si observem atentament entremig de les bràctees, trobarem una pell fina i transparent, que es pot treure fàcilment. És com un tel, humit, aparentment homogeni quan s’observa a ull nu.
Però si observem aquesta pell fina al microscopi veurem que és formada per polígons allargats, els uns enganxats als altres com les rajoles del terra: són les cèl·lules. Si hi tirem una gota de colorant (per exemple una solució iodurada diluïda) aconseguirem veure dins les cèl·lules uns corpuscles petits: són els nuclis.
El nostre cos també amaga aquestes estructures, invisibles a primer cop d’ull. La sang que raja quan ens punxem un dit té l’aspecte d’un líquid vermell, però el seu examen al microscopi revela la presència de petits disquets rodons: són els glòbuls vermells. Aquestes paraules que ara llegiu són visualitzades, conduïdes al cervell i enteses gràcies a les cèl·lules nervioses.
Al món hi ha milions d’espècies diferents d’éssers vius, des del protozou microscòpic fins a l’enorme balena, des dels fongs a l’espècie humana, des del til·ler a la medusa. Són molt diferents quant a dimensions, formes i hàbits, tot i que, tret d’alguns organismes d’estructura molt simple (els virus), tots els éssers que viuen o han viscut a la Terra s’han format a partir d’una unitat fonamental: la cèl·lula.
La cèl·lula, unitat fonamental de la vida
Les cèl·lules de tots els éssers vius són bàsicament semblants, és a dir, han resolt d’una manera anàloga les principals qüestions vitals: reproduir-se, créixer i intercanviar matèria i energia amb el medi circumdant. Hi ha organismes, com els virus i els bacteriòfags (virus capaços d’infectar els bacteris), que són més simples que la cèl·lula. A la pràctica només són capaços de reproduir-se i, per tant, no poden ser considerats autònoms. Per a viure han d’allotjar-se a l’interior d’una cèl·lula hoste. Com a exemple tenim els que provoquen el refredat, la grip o el xarampió.
Hi ha éssers vius formats per una sola cèl·lula i d’altres formats per moltes cèl·lules. Els éssers unicel·lulars proliferen en grans quantitats (centenars de milions) en tots els ambients, però són invisibles a ull nu. Fins i tot a la superfície més neta, si s’examina a fons, es descobrirà la seva presència. Molts d’aquests organismes també viuen dins nostre: n’hi ha de perjudicials, d’innocus i fins i tot d’útils. A l’intestí humà, per exemple, hi ha bacteris capaços de produir la vitamina K. Les vaques, els cavalls i els herbívors en general també "allotgen" en l’aparell digestiu hostes microscòpics que els resulten indispensables per a poder digerir la cel·lulosa.
Els organismes formats per una sola cèl·lula es divideixen en dos grans grups o regnes: moners i protoctists. El regne dels moners comprèn els bacteris i els cianobacteris (o algues blaves), que són formats per una cèl·lula primitiva, i per això també s’anomenen procariotes (mot derivat del grec que significa "abans del nucli"). El seu material genètic no s’organitza a l’interior d’un nucli. Són els organismes més antics i els més nombrosos del món. Els protoctists, en canvi, són formats per una única cèl·lula (o com a màxim unes quantes) més complexa anomenada eucariota (terme que deriva del grec i significa "nucli ben fet"). Són protoctists, per exemple, els protozous.
Els altres éssers vius també són constituïts de cèl·lules eucariotes, generalment nombroses (en l’espècie humana unes 1014, o sigui cent bilions), diversament especialitzades i organitzades en teixits i òrgans. Els organismes pluricel·lulars es divideixen en els següents regnes: fongs, vegetals i animals.
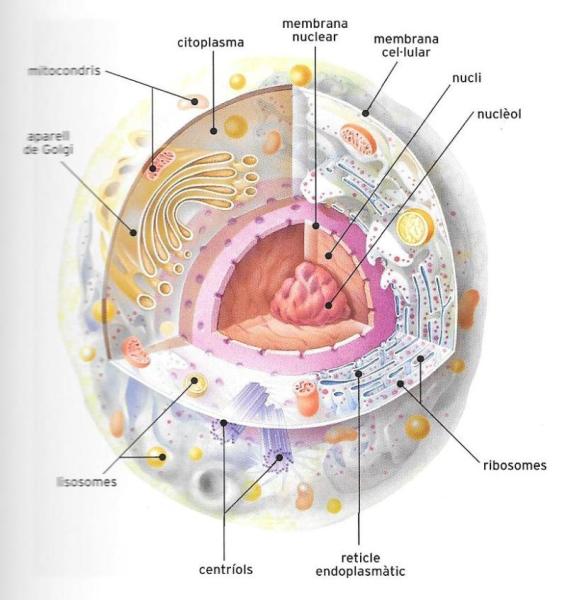
Estructura d'una cèl·lula eucariota tipus, no especialitzada, en la qual s'han representat els elements morfològics més generals.
ECSA
Tot organisme pluricel·lular depèn de les seves pròpies cèl·lules per a viure, cadascuna de les quals creix, consumeix i produeix energia, es multiplica i, alhora, contribueix a la vida del conjunt de l’organisme. L’espècie humana prové d’una única cèl·lula, el zigot, fruit de la unió de l’espermatozoide del pare i l’òvul de la mare. Durant el desenvolupament anterior al naixement, aquesta única cèl·lula es reprodueix i dóna lloc a dues cèl·lules filles. Aquestes cèl·lules també es reprodueixen, originant un procés que continua fins a la formació dels milers de milions de cèl·lules que componen el cos humà en el moment del naixement. Totes aquestes cèl·lules pateixen un fenomen d’especialització: algunes, que esdevenen capaces d’estirar-se i arronsar-se, formen els músculs; d’altres acumulen substàncies minerals i formen els ossos, estructures robustes i elàstiques; n’hi ha que constitueixen llargs filaments a través dels quals circulen els estímuls nerviosos, etc. És el conjunt coordinat de totes les cèl·lules allò que fa funcionar el nostre cos i li dóna vida.
Com són de grans, les cèl·lules?
Les cèl·lules són molt petites, tan petites que no són visibles a ull nu i ni tan sols amb la lupa; cal recórrer al microscopi. Per a mesurar-les no es pot utilitzar el mil·límetre com a unitat de longitud, sinó la mil·lèsima part, és a dir, el micròmetre o micra, que s’indica amb el símbol µm. Hi ha parts de la cèl·lula per a les quals fins i tot el micròmetre és massa gran, aleshores es recorre al nanòmetre (símbol nm) o mil·limicra, que és la mil·lèsima part del micròmetre. La majoria de les cèl·lules eucariotes tenen un diàmetre que oscil·la entre 10 i 30 µm, però n’hi ha de més petites. El cerebel conté les cèl·lules més petites del cos humà, que fan tan sols 4 µm de diàmetre. Però també hi ha cèl·lules molt més grans. En els animals, per exemple, les cèl·lules nervioses amb les seves prolongacions assoleixen dimensions considerables. Excepcionalment, fins i tot hi ha cèl·lules que són visibles a ull nu; només cal pensar en els òvuls de molts animals (els ocells i els rèptils), que de vegades són molt voluminosos perquè contenen substàncies de reserva acumulades (els ous de les gallines fan de 5 a 7 cm de llargada i pesen de 45 a 70 g).
De les cèl·lules visibles a ull nu també cal destacar l’acetabulària. Quan s’exploren els esculls marins amb les ulleres és fàcil veure-hi matolls d’algues que recorden petites margarides; són justament les acetabulàries, el cos de les quals és format per una sola cèl·lula gegantina, d’entre 2 i 5 cm de llarg, diferenciada en capçada, tija i peu.
De tota manera, una cèl·lula normalment no pot superar un volum determinat. En primer lloc, perquè el nucli (que, com veurem, dirigeix tota l’activitat cel·lular) no pot controlar una cèl·lula massa voluminosa (algunes cèl·lules grans tenen diversos nuclis idèntics, per exemple el protozou parameci).
La segona causa que limita les dimensions d’una cèl·lula és el fet que totes les substàncies que hi entren i en surten i que, per tant, en determinen la vida, han de travessar la membrana que la delimita. Per exemple, hi entren oxigen i aliments, i en surten substàncies de rebuig i altres materials. Ara bé, mentre la cèl·lula creix, l’augment de superfície en relació amb el del volum és menor, ja que l’augment superficial és proporcional al quadrat de les dimensions lineals, mentre que el del volum ho és al cub. Així, les cèl·lules més petites tindran una superfície proporcionalment més àmplia per a intercanviar substàncies amb el medi extern. Un cop assolida una certa dimensió, l’extensió de la membrana serà insuficient per a recobrir tot el volum d’una manera adequada. El volum de les cèl·lules és, doncs, força definit i depèn principalment de les seves funcions: les cèl·lules del fetge d’un ratolí són tan grans com les del fetge d’un cavall. Allò que determina la diferència entre els fetges de tots dos animals, i en general dels seus cossos, és el nombre de cèl·lules. Les cèl·lules nervioses, que sovint són proporcionals al volum de l’animal, són l’excepció d’aquesta norma.
Forma i estructura general de les cèl·lules
Una cèl·lula jove, encara no especialitzada, té tendència a adoptar la forma esfèrica. En general, però, a mesura que la cèl·lula es diferencia per a desenvolupar una funció determinada, adopta una forma característica, que és conseqüència tant de la seva estructura interna com de la pressió de les cèl·lules veïnes. Així, una cèl·lula de la pell, aïllada i introduïda en un líquid adequat, esdevé esfèrica, mentre que, en el teixit, el contacte amb les altres cèl·lules la fa esdevenir polièdrica. Els protoctists, com els protozous, presenten formes molt diverses tot i constar d’una sola cèl·lula, car han d’assumir totes les funcions existencials (moure’s, defensar-se, etc.). Per això aquestes cèl·lules tenen formes enormement complexes. En el fons, però, responen al pla bàsic de tota cèl·lula.
Malgrat la gran varietat existent d’éssers vius, totes les cèl·lules presenten una sèrie d’estructures iguals, fins al punt que es pot caracteritzar una cèl·lula tipus. Aquestes estructures són les següents:
1) la membrana, que separa la cèl·lula del medi extern i en permet i regula els intercanvis d’oxigen, aigua, sals minerals, substàncies nutritives, etc. Pot tenir un recobriment exterior, que en les cèl·lules vegetals és una paret força rígida constituïda sobretot de cel·lulosa, en la qual es poden incorporar altres substàncies, com la lignina;
2) el citoplasma, amb diversos aparells i òrgans, on es desenvolupen les activitats del metabolisme cel·lular;
3) el nucli, que en les cèl·lules eucariotes és recobert d’una membrana pròpia. Conté la informació per al desenvolupament i la duplicació cel·lular: totes les cèl·lules capaces de reproduir-se tenen com a mínim un nucli.
Les cèl·lules procariotes (els bacteris i els cianobacteris) estan mancades de membrana nuclear. Els virus, com ja hem remarcat, són dotats d’un sistema que els permet reproduir-se, però, en canvi, no disposen de metabolisme propi. Són paràsits obligats i no es consideren veritables cèl·lules.
La membrana cel·lular
Totes les cèl·lules viuen en un medi fluid. Per als protozous pot ser l’aigua; per a les cèl·lules dels organismes pluricel·lulars són els líquids corporals (la sang, la limfa i el líquid intersticial). No hi ha cap cèl·lula que pugui viure en un medi sec. L’interior de la cèl·lula també és fluid, i és de vital importància que mantingui una composició determinada, diferent de la del medi circumdant; d’això se n’encarrega la membrana cel·lular.
Funcions de la membrana cel·lular
La membrana cel·lular determina i manté les diferències entre el líquid de l’interior de la cèl·lula i el de l’exterior, bo i controlant quines substàncies en surten, quines hi entren i en quina quantitat. Ens podem imaginar la membrana cel·lular com les muralles d’un castell: han de ser robustes i dotades de sistemes de defensa però, al mateix temps, han de permetre l’entrada i la sortida. En efecte, a l’interior del castell es realitzen moltes activitats, per a les quals es requereixen materials procedents de l’exterior en quantitats precises. El ferrer necessita ferro per a forjar, però si en rebés sense parar, més del que li cal, tard o d’hora hi hauria ferro arreu del castell: ferro a les quadres, a les cuines, als patis, als pous... En definitiva, totes les activitats quedarien interrompudes. D’altra banda, si no n’hi arribés prou, els cavalls no podrien ser ferrats, ni els carros reparats, etc.; totes les activitats del castell se’n ressentirien. Al mateix temps, cal treure a fora totes les deixalles. El ferrer endreçat escombra sovint el seu taller, i totes les deixalles són apartades del castell.
A més del ferrer hi ha el forner, el llenyataire, etc., tots amb problemes semblants: han de produir objectes determinats, necessaris per a la vida del castell; per tant, han de rebre la quantitat necessària de material i en el moment més oportú. A més, els cal desfer-se de les deixalles i potser enviar productes a algun altre castell.
Les portes del castell són custodiades per guàrdies que tenen ordres ben concretes: alguns materials passen sense problemes, d’altres són rebutjats, i n’hi ha que només passen si un acompanyant conegut en respon. Tant les ordres com els guàrdies canvien segons les necessitats del castell.
De la mateixa manera, la membrana cel·lular deixa passar o no determinades substàncies, en funció de les necessitats de la cèl·lula. Els "guàrdies" són les molècules que la constitueixen. Allò que regula l’intercanvi de materials entre la cèl·lula i el medi és el tipus de molècules, controlat bàsicament pel nucli (l’encarregat de la jurisdicció del castell). També es pot pensar que els diferents tallers del castell són els diversos orgànuls del citoplasma (els mitocondris, els lisosomes, etc.), dels qual parlarem ben aviat, cadascun especialitzat en una activitat concreta.
Estructura de la membrana cel·lular
La membrana cel·lular fa uns 8 nm de gruix. Per tant, només es pot veure amb el poderós augment que proporciona el microscopi electrònic. Constitueix una protecció robusta i versàtil i és formada per unes molècules especials: els fosfolípids (constituïts per lípids, és a dir greixos, per fòsfor i per nitrogen). Aquestes molècules presenten una particularitat: una de les seves extremitats és atreta per l’aigua (hidròfila), però l’altra no (hidròfoba). En la membrana cel·lular es disposen en una doble capa, ordenades de manera que totes les extremitats hidròfiles es dirigeixen cap al líquid. Així, a una banda queda el citoplasma i a l’altra el líquid extracel·lular. Les extremitats hidròfobes, en canvi, queden encarades; el conjunt pot recordar un entrepà.
A més dels fosfolípids, també són presents a la membrana moltes proteïnes. Algunes són estructurals, és a dir, formen part de la mateixa membrana; d’altres, els enzims, serveixen per a afavorir certes reaccions químiques; i n’hi ha d’altres que són "transportadores", acompanyen determinades substàncies a través de la membrana, com veurem de seguida.
A la cara externa de la membrana hi ha cadenes de glúcids (sucres) que tenen la funció, sobretot, de "reconèixer" les molècules presents a l’interior de la cèl·lula i d’unir les cèl·lules adjacents.
A la cèl·lula hi ha moltes membranes més. El nucli i altres components cel·lulars com els mitocondris i els cloroplasts (dels quals parlarem més endavant) posseeixen un embolcall propi. Totes les membranes tenen l’estructura bàsica d’una doble capa de fosfolípids amb proteïnes; ara bé, la composició química varia i, per tant, el comportament de la membrana és diferent. En efecte, les substàncies que entren i surten del nucli són en bona part diferents de les que entren i surten d’un mitocondri (de la mateixa manera que són diferents la primera matèria i els productes d’un forner i d’un llenyataire).
Les molècules que componen les membranes es renoven contínuament, fins i tot quan la cèl·lula ha completat el seu creixement: la seva integritat perfecta és d’una importància vital.
Particularitats de la membrana
El nostre cos, com el de tots els animals pluricel·lulars, funciona gràcies a la coordinació de l’activitat dels milers de milions de cèl·lules que el componen. Així doncs, d’alguna manera, es comuniquen entre elles. Aquesta comunicació es pot donar a través de les substàncies transportades per la sang, com les hormones, o mitjançant el sistema nerviós; però també pot ser directa, de cèl·lula a cèl·lula.
En efecte, moltes cèl·lules tenen uns canals minúsculs que connecten la seva membrana amb la de les cèl·lules contigües, i això permet el pas de certes molècules. Així, cada cèl·lula és "informada" de la situació de les seves veïnes. En el teixit nerviós i en el muscular, aquest tipus d’unió permet que l’estímul es propagui amb molta facilitat a totes les cèl·lules situades en una certa zona, que així es comporten d’una manera uniforme, com una unitat funcional.
A més, quasi totes les cèl·lules animals tenen damunt la membrana un recobriment de glicoproteïnes (proteïnes unides a glúcids) que és fonamental perquè una cèl·lula i l’altra es reconeguin, cosa que fa possible els intercanvis regulars.
Les microvellositats són una altra diferenciació de la membrana cel·lular. Es tracta de prolongacions més o menys gruixudes i regulars, que augmenten enormement la superfície de la cèl·lula. Són especialment evidents en les cèl·lules de l’epiteli dels túbuls renals i en les de l’epiteli intestinal, del qual augmenten la capacitat d’absorció. En efecte, és a través d’aquestes cèl·lules que les substàncies nutritives digerides passen a la sang, des d’on són distribuïdes per tot el cos. Les microvellositats es poden allargar i encongir, però no es poden moure; al contrari dels cilis, apèndixs del citoplasma, dels quals es tractarà en "El citoplasma".
Com s’entra a la cèl·lula i com se’n surt
La membrana cel·lular és semipermeable, és a dir pot deixar passar certes substàncies però no d’altres. Aquesta permeabilitat selectiva és allò que manté la composició del citoplasma en els seus precisos termes.
El pas a través de la membrana (de l’interior a l’exterior i viceversa) es dóna bàsicament de tres maneres: 1) per difusió, 2) per transport actiu i 3) per endocitosi o exocitosi.
1) Difusió. El mecanisme de pas per difusió tendeix a equilibrar la quantitat d’una certa substància a l’interior i a l’exterior de la cèl·lula. Si, per exemple, a l’exterior de la cèl·lula hi ha una concentració d’oxigen més gran que a l’interior, l’oxigen entra dins la cèl·lula. Això s’esdevé més ràpidament com més gran és la diferència de concentració (gradient). Quan s’assoleix l’equilibri l’oxigen continua travessant la membrana, però ara ho fa en ambdós sentits i en la mateixa quantitat. Les molècules petites o solubles en els greixos presents en la membrana passen per difusió simple. L’aigua també passa per difusió i, en aquest cas, es parla d’osmosi.
Algunes molècules no tenen autonomia per a travessar la membrana cel·lular. Aleshores intervenen proteïnes especials (anomenades transportadores o també permeases) que s’uneixen a les molècules i les "acompanyen". Hi ha una proteïna "transportadora" per a cada molècula. La seva presència o absència contribueix a determinar les diferències entre les cèl·lules.
2) Transport actiu. La difusió, que tendeix a equilibrar la concentració d’algunes substàncies a l’interior i a l’exterior de la cèl·lula, implicaria la uniformitat de determinades molècules. Però en molts casos no és així. Per exemple, en el citoplasma hi ha molt més potassi (K) que a l’exterior de la cèl·lula, on és més gran la concentració de sodi (Na). La membrana manté aquesta diferència per mitjà del "transport actiu", mecanisme que actua contra el gradient de concentració i, per tant, consumeix energia.
3) Endocitosi i exocitosi. Tots els glòbuls blancs (leucòcits) es mouen per la sang emetent prolongacions de citoplasma (pseudòpodes); quan entren en contacte amb un bacteri, l’embolcallen en una mena d’abraçada. Així, el bacteri queda atrapat en un sac de glòbul blanc. Alguns sucs digestius destrueixen el cos del bacteri i el seu contingut, que és absorbit pel citoplasma del leucòcit. Aquest mecanisme s’anomena endocitosi (és a dir, introducció en la cèl·lula). Si el material englobat és constituït per partícules sòlides prou voluminoses, com en el cas del bacteri acabat de descriure, es parla de fagocitosi (del grec "fago", menjo). En canvi, si es tracta de gotetes de líquid, l’endocitosi rep el nom de pinocitosi (del terme grec "pino", que significa bec).
Totes les cèl·lules eucariotes són capaces d’englobar al seu interior corpuscles o líquids per endocitosi. El material és "reconegut" per la membrana cel·lular, que forma una vesícula al seu voltant. Aquesta vesícula es desprèn de la membrana i s’allibera al citoplasma, on, en alguns casos, els enzims digestius la dissoldran. És d’aquesta manera com "mengen" molts protozous com les amebes, o com els glòbuls blancs i els macròfags ens protegeixen de les partícules de pols de l’aire o dels microorganismes que envaeixen el nostre organisme i en destrueixen les cèl·lules velles (vegeu "El cos humà, la salut i la malaltia").
En altres casos, la vesícula d’endocitosi travessa el citoplasma i per mitjà d’un mecanisme contrari —l’exocitosi— és expulsada a l’altre costat. És així com, per exemple, el material nutritiu travessa les cèl·lules de l’epiteli intestinal per a passar de l’intestí a la sang.
Hi ha altres vesícules que es queden dins la cèl·lula, plenes de material de reserva. Això és el que passa, per exemple, en els òvuls. Alguns virus, bacteris i protozous que han de viure dins la cèl·lula per a reproduir-se, s’aprofiten d’aquest mecanisme d’endocitosi de la cèl·lula per entrar-hi i desenvolupar-hi la seva vida paràsita.
El citoplasma
A través de la membrana arriben al citoplasma moltes substàncies, en particular aliments i oxigen, i allí són transformades gràcies a l’oxigen. La cèl·lula obté dels aliments l’energia necessària per a desenvolupar les seves múltiples activitats.
El citoplasma té una estructura molt complexa i molt organitzada. Bàsicament és format per: aigua, on hi ha nombroses substàncies dissoltes o en suspensió que li confereixen un aspecte dens i viscós; un conjunt de membranes internes, unides entre elles i diversament diferenciades; una xarxa de túbuls i filaments, que constitueix l’esquelet de la cèl·lula, i diversos orgànuls amb funcions específiques.
Les cèl·lules vegetals tenen estructures i òrgans específics que tractarem més endavant, a "La cèl·lula vegetal". Aquí il·lustrem els òrgans comuns a totes les cèl·lules.
Les membranes internes
El microscopi electrònic ha fet visible, gràcies al seu poderós augment, una vasta xarxa de membranes a l’interior de la cèl·lula, on abans es pensava que no hi havia estructures. Aquestes membranes formen l’embolcall nuclear, el reticle endoplasmàtic i l’aparell de Golgi.
El reticle endoplasmàtic, present en totes les cèl·lules, apareix particularment desenvolupat en aquelles amb una producció intensa de proteïnes, com per exemple les cèl·lules glandulars. Juntament amb el citoesquelet, contribueix a donar forma i una relativa rigidesa a la cèl·lula, i regula els intercanvis entre el citoplasma i l’exterior. El reticle endoplasmàtic és format per membranes que, replegades, delimiten una mena de "cisternes". En algunes zones tenen molts ribosomes (petits orgànuls sobre els quals ja insistirem), mentre que en d’altres no n’hi ha. En el primer cas es parla de reticle endoplasmàtic rugós, i en el segon, de reticle endoplasmàtic llis.
En el reticle endoplasmàtic llis s’han observat diverses activitats: algunes substàncies tòxiques, com per exemple plaguicides, additius químics, hipnòtics, se solubilitzen i així poden passar a la sang i ser eliminades mitjançant l’excreció. També s’hi ha observat la formació de molècules de greixos i de glicogen, substàncies de reserva per a l’organisme.
Els ribosomes, presents en el reticle endoplasmàtic granular o escampats pel citoplasma, poden estar disposats en cadenes més o menys llargues i en ells s’esdevé la síntesi de les proteïnes. Cada cèl·lula de l’organisme produeix proteïnes específiques que determinen tant la naturalesa de la cèl·lula com les reaccions químiques que hi tenen lloc. És sobretot el tipus de proteïnes el que fa que una cèl·lula muscular sigui diferent d’una cèl·lula de la pell i que un cavall sigui diferent d’una tortuga.
Una part del sistema de membranes internes té un aspecte diferent del del reticle endoplasmàtic. És formada per una sèrie de sàculs arrenglerats, connectats a vesícules petites i grans (vacúols), i rep el nom d’aparell o complex de Golgi (del nom de Camillo Golgi, que el va descobrir el 1898). L’aparell de Golgi serveix per a emmagatzemar i distribuir les substàncies produïdes per l’activitat cel·lular. A més, hi té lloc la formació d’algunes molècules complexes, com les glicoproteïnes esmentades abans, que es troben a la membrana cel·lular, les lipoproteïnes (formades per proteïnes i greixos) i els polisacàrids. Quan cal, aquestes substàncies són menades fora de la cèl·lula per exocitosi: els vacúols es desplacen cap a la perifèria de la cèl·lula, s’adhereixen a la membrana plasmàtica i s’obren a l’exterior, on alliberen el contingut.
Dels vacúols de l’aparell de Golgi s’originen els lisosomes, vesícules amb funcions digestives (vegeu "Els lisosomes, els escombriaires cel·lulars"). Efectivament, contenen nombrosos enzims que transformen les proteïnes, els glúcids, els greixos i els àcids nucleics en molècules més simples.
Els lisosomes, els escombriaires cel·lulars
Els lisosomes (del grec "soma", cos, i "liso", destruir; o sigui, cossos destructors) són orgànuls delimitats per una membrana i que contenen substàncies (en concret, enzims hidrolítics) capaces de degradar —o sigui, de transformar en molècules simples— pràcticament tots els tipus de molècules orgàniques complexes: les proteïnes, els polisacàrids, els àcids nucleics i els lípids. Per aquest motiu la missió fonamental dels lisosomes és ocupar-se de la digestió cel·lular. Intervenen en molts moments al llarg de la vida d’un organisme o d’una cèl·lula. Per exemple, poden reabsorbir parts d’òrgans o òrgans sencers. Així, l’úter d’una dona, després del part, es redueix de 2 kg a 50 g en 9 dies per l’acció dels lisosomes. Una granota acabada de néixer és molt diferent dels seus pares, és un capgròs amb cua i brànquies. A mesura que va creixent es va transformant a poc a poc en una granoteta: les brànquies desapareixen, comencen a sortir les potes i després desapareix la cua; els lisosomes en "digereixen" els teixits.
Els lisosomes també són els encarregats de la destrucció química del material introduït en la cèl·lula per endocitosi: es fonen amb la vesícula d’endocitosi i en digereixen el contingut. Constitueixen l’aparell digestiu dels protozous, i destrueixen els bacteris continguts en els glòbuls blancs de la sang. En general, aquestes vesícules digestives particulars són més nombroses en algunes cèl·lules com, precisament, els glòbuls blancs o leucòcits, els macròfags, grans cèl·lules mòbils escampades en diversos centres de l’organisme, o bé els osteoclasts, cèl·lules encarregades de la destrucció del teixit ossi vell, que es renova contínuament.
Si els enzims dels lisosomes sortissin al citoplasma, el destruirien. És el que passa en algunes malalties, com la gota (causada per l’acumulació de substàncies anomenades urats en les articulacions), la silicosi (produïda per la inhalació de sílice, component del vidre) i l’asbestosi (provocada per la inhalació d’amiant). Aquests materials provoquen l’alliberament d’enzims per part dels lisosomes, amb la consegüent inflamació dels teixits.
L'"esquelet" de la cèl·lula
A l’interior del citoplasma hi ha una xarxa de túbuls, filaments i, a vegades, trabècules (estructures en forma de bastonets), visibles al microscopi electrònic. Constitueixen el citoesquelet, que manté la forma de la cèl·lula i la posició dels diversos orgànuls.
No és una estructura fixa ni rígida, sinó que es modifica, contribueix al moviment cel·lular i regula els desplaçaments al seu interior.
Els filaments són constituïts per proteïnes filamentoses, entre les quals hi ha, per exemple, l’actina i la miosina, que, en lliscar l’una sobre l’altra, escurcen les cèl·lules musculars.
Els túbuls, compostos també per proteïnes globulars, formen la paret dels cilindres buits, llargs i prims. A més de ser una part del citoesquelet veritable, també formen algunes estructures específiques: els cilis, els flagels, l’aparell de l’astrosfera i el fus mitòtic.
Cilis i flagels
Els cilis i els flagels són apèndixs prims del citoplasma dotats de moviment. Si són pocs i llargs s’anomenen flagels, com per exemple en els protozous flagel·lats. En canvi, si són curts i nombrosos, adopten el nom de cilis, com en els protozous ciliats o en les cèl·lules de la superfície interna d’alguns trams de les vies reproductores i de les respiratòries. Són presents en quasi tots els organismes eucariotes, per bé que no en totes les seves cèl·lules. En són una excepció les plantes amb flors, en les quals no hi ha cèl·lules ciliades. Per a molts protozous en constitueixen el sistema de locomoció.
L’espermatozoide, cèl·lula germinal masculina, se serveix de la cua flagel·lada per a moure’s i, juntament amb milions d’altres espermatozoides, nedar ràpidament cap a l’òvul per a fecundar-lo. L’òvul, cèl·lula germinal femenina que viatja cap a l’úter, és mogut pels cilis que recobreixen les parets de les trompes. Els cilis de l’interior dels bronquis transporten en sentit ascendent, amb el seu moviment sincronitzat, les substàncies estranyes que podem haver inhalat amb l’oxigen, així es poden remoure i expulsar la brutícia, la pols i els mocs. Els cilis i els flagels són formats per estructures tubulars disposades de manera regular: un parell de túbuls central, circumdat de nou parells col·locats ordenadament al seu voltant formant un cercle. El moviment dels túbuls, causat per les característiques de les proteïnes que els formen i per la seva estructura, provoca el moviment del cili i el flagel.
La centrosfera
Més o menys al centre de moltes cèl·lules eucariotes (excepte en les plantes amb flors) hi ha una zona particularment estructurada, anomenada centrosfera. Aquesta zona es modifica profundament durant la divisió cel·lular.
Quan la cèl·lula no s’està dividint, és formada per dos corpuscles (anomenats centríols), envoltats d’una corona de filaments (anomenats àsters) o astrosfera. Els centríols i l’astrosfera són formats per microtúbuls.
Durant la divisió cel·lular els dos centríols s’allunyen mútuament fins a situar-se als pols oposats de la cèl·lula, envoltats per l’astrosfera en tot moment. L’espai que els separa és ocupat per un feix de fibres formades de microtúbuls. Així es constitueix el fus mitòtic, que té un paper fonamental en la reproducció cel·lular.
Els mitocondris
Quan pugem a una muntanya, nedem molta estona, o fem una cursa, se’ns accelera la respiració, el cor batega més de pressa i tenim gana. Per què? Les nostres cèl·lules musculars, sotmeses a un esforç, requereixen oxigen i nutrients. Llavors, respirem més de pressa i el cor ens batega més ràpid perquè la sang els faci arribar més oxigen, i mengem més perquè els arribin més substàncies nutritives.
Energia per a viure
A l’interior del citoplasma de totes les cèl·lules eucariotes hi ha uns corpuscles anomenats mitocondris (del grec "mitos", filament, i "condros", raïm). Amb l’ajut de l’oxigen obtenen de les substàncies nutritives l’energia necessària per a totes les activitats cel·lulars. Els mitocondris són els encarregats de produir la calor corporal. Aquesta calor s’ha de mantenir constant sigui quina sigui la temperatura exterior, l’energia necessària per a la contracció muscular, el moviment dels cilis de les cèl·lules, el "trànsit" de les substàncies a través de les membranes cel·lulars i el pas dels estímuls nerviosos.
L’energia necessària per a la síntesi de molècules noves, per exemple la formació de proteïnes noves a partir de nombrosos aminoàcids, sempre prové dels mitocondris.
Els mitocondris també proporcionen l’energia que produeix la llum de les lluernes i la descàrrega elèctrica de les tremoloses. Els exemples són, doncs, innombrables.
Els mitocondris tenen gairebé sempre forma ovoïdal i són més nombrosos com més intensa és l’activitat de la cèl·lula; poden modificar-se i desplaçar-se pel citoplasma segons les exigències metabòliques. Són formats per dues membranes, una d’externa, llisa, i una d’interna, replegada en diverses "crestes mitocondrials". Així, queden delimitats dos espais: un entre les membranes, anomenat cambra mitocondrial externa, i un altre d’intern, anomenat cambra mitocondrial interna, que conté un líquid dens: la matriu mitocondrial. Totes les parts del mitocondri són químicament diferents i assumeixen distintes funcions.
Oxigen, aliments i energia
Les molècules que constitueixen els nostres aliments contenen enllaços químics que, en trencar-se, alliberen energia. La glucosa produeix energia en una sèrie de reaccions que es poden resumir així: glucosa + oxigen → diòxid de carboni + aigua + energia; o bé:
C6H12O6 + 6 O2 → 6 CO2 + 6 H2O + 686 kcal/mol
La glucosa és produïda per les plantes per mitjà de la fotosíntesi clorofíl·lica (descrita a "El món dels vegetals" ). En aquest procés l’energia del Sol és "capturada" (gràcies a la clorofil·la) i emprada per a formar glucosa a partir de l’aigua i el diòxid de carboni. És a dir: aigua + diòxid de carboni + energia → glucosa + oxigen; o bé:
6 H2O + 6 CO2 + energia → C6H12O6 + 6 O2
Tal com es pot veure, es tracta de la reacció contrària a la que s’ha escrit més amunt. Així doncs, nosaltres, com tots els éssers vius dotats de mitocondris —és a dir, tots els eucariotes—, utilitzem en darrera instància l’energia del Sol per a viure, energia que forma part dels enllaços químics de la glucosa produïda per les plantes i de les quals forma part. A través de la cadena alimentària la glucosa arriba a tots els éssers vius: nosaltres ens nodrim directament de vegetals o bé d’animals que s’han nodrit de vegetals. A més dels glúcids, els greixos i les proteïnes també produeixen energia, intervenen en les reaccions que tenen lloc en els mitocondris (vegeu també "La nutrició").
El 40% de l’energia continguda en la glucosa és a disposició de la cèl·lula. L’eficàcia és notable si es té en compte que el motor d’un cotxe solament aprofita el 25% de l’energia continguda en la benzina, mentre que el 75% restant es perd en forma de calor.
L’energia obtinguda de la glucosa serveix per a formar una molècula determinada, l’ATP (trifosfat d’adenosina), que és cedit al metabolisme cel·lular d’una manera controlada i funcional.
L’ATP és format per una base nitrogenada, l’adenina, per un sucre, la ribosa, i per tres grups fosfat.
Els enllaços que uneixen els grups fosfat es trenquen fàcilment i alliberen molta energia. Normalment només es trenca un d’aquests enllaços i l’ATP esdevé ADP (difosfat d’adenosina) per la pèrdua d’un fosfat inorgànic (Pi) que allibera 7 kcal per mol aproximadament. De la mateixa manera, l’ADP pot tornar a convertir-se en ATP i proporcionar-li 7 kcal:
ATP ⇄ ADP + 7 kcal + Pi
L’energia necessària per a formar ATP a partir d’ADP la proporciona l’oxidació de la glucosa (vegeu "Com es forma l’ATP"). L’ATP pot ser utilitzat per la cèl·lula bàsicament de dues maneres:
a) essent transformat en ADP per determinats enzims anomenats ATPases. L’energia obtinguda és utilitzada immediatament, per exemple, per a produir calor;
b) perdent un grup fosfat que és transferit a una altra molècula per determinats enzims anomenats cinases i s’hi adhereix (fosforilació). La molècula carregada d’energia d’aquesta manera participa en una altra reacció. És el que s’esdevé, per exemple, en la formació de la sacarosa (un sucre disacàrid) a partir de la glucosa i la fructosa (dos sucres monosacàrids). Els monosacàrids són fosforilats i a continuació es forma el disacàrid:
ATP + glucosa → glucosa-fosfat + ADP;
ATP + fructosa → fructosa-fosfat + ADP;
glucosa-fosfat + fructosa-fosfat → sacarosa + 2 Pi.
Com es forma l’ATP
L’energia per a formar molècules d’ATP prové de l’oxidació de la glucosa, que comença en el citoplasma, amb un procés anomenat glicòlisi anaeròbica, perquè no requereix oxigen. Una molècula de glucosa, que té 6 àtoms de carboni, és escindida en dues molècules d’un compost més petit, l’àcid pirúvic, amb 3 àtoms de carboni. L’energia alliberada serveix per a formar dues molècules d’ATP:
C6H12O6 + 2ADP + 2Pi → 2C3H6O3 + 2ATP
L’oxidació de la glucosa continua a l’interior dels mitocondris, on se cedeix la major part de l’energia, amb la formació de 36 molècules més d’ATP (en condicions òptimes).
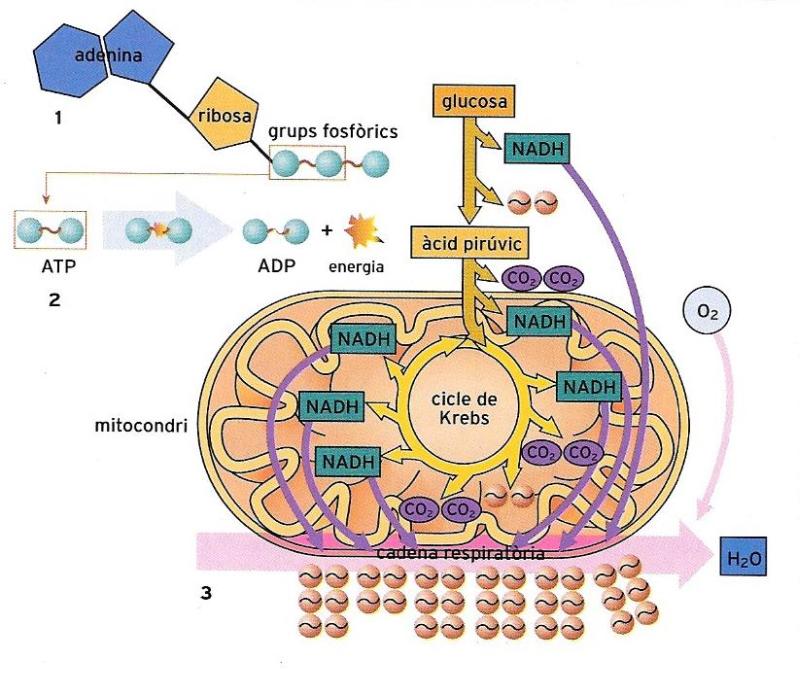
Representació simplificada d'una molècula d'ATP (1) i de l'alliberament d'energia per ruptura d'un dels enllaços fosfat, altament energètics (2). A baix (3),transformació de la glucosa a través de la glicòlisi i la cadena respiratòria a l'interior del mitocondri, amb la intervenció dels transportadors FAD (en estat reduït FAD+ o FADH) i NAD (NAD+ o NADH). En totes les oxidacions biològiques, els carburants metabòlics com ara la glucosa actuen de donadors d'electrons, mentre que els nucleòtids NAD+ i FAD actuen d'acceptors d'electrons.
ECSA
Les reaccions que tenen lloc en els mitocondris reben el nom de respiració cel·lular i requereixen oxigen. La respiració inclou dos processos: el cicle de Krebs, que té lloc a la matriu mitocondrial, i la fosforilació oxidativa, que té lloc a les crestes mitocondrials. En aquestes fases tenen un paper molt important certes molècules (NAD+ i FAD+), encarregades del transport de l’hidrogen i l’energia obtinguts en el procés de destrucció de la glucosa fins al seu destí definitiu: la formació d’aigua i d’ATP. El balanç global és, com a màxim, de 38 molècules d’ATP per cada molècula de glucosa. Quina part de l’energia de la glucosa s’ha aconseguit aprofitar en aquestes 38 molècules? Per cada molècula de glucosa s’alliberen 686 kcal. La síntesi d’una molècula d’ATP requereix unes 7 kcal. Per tant, són 266 (7 x 38) les kcal emmagatzemades per l’ATP, mentre que l’energia restant es dissipa en forma de calor.
Resumim en una fórmula el que acabem de dir:
C6H12O6 + 6O2 + 38ADP + 38Pi → 6CO2 + 6H2O + 38ATP + 440 kcal
Les molècules d’ATP són conduïdes fora dels mitocondris, alhora que s’hi introdueix el mateix nombre de molècules d’ADP.
Quan no hi ha oxigen, no hi pot haver respiració en els mitocondris. Això passa, per exemple, durant una intensa activitat muscular; la respiració augmenta de ritme i el cor batega més de pressa. Certament, cal molt oxigen per a produir l’ATP necessari per a contreure els músculs. Però sovint l’oxigen és insuficient, aleshores les cèl·lules continuen transformant glucosa en àcid pirúvic, reacció que té lloc en el citoplasma sense necessitat d’oxigen, i produeixen 2 molècules d’ATP per cada molècula de glucosa. Ara bé, com que no hi ha prou oxigen, l’àcid pirúvic no s’oxida, sinó que es transforma en àcid làctic. L’acumulació d’àcid làctic és el que produeix la sensació de cansament muscular (el cruiximent). Després, quan reposem, l’àcid làctic es torna a transformar en àcid pirúvic i posteriorment en glucosa:
glucosa ⇄ àcid pirúvic ⇄ àcid làctic
Les cèl·lules del llevat, en canvi, quan no hi ha oxigen transformen la glucosa del raïm en àcid pirúvic, que al seu torn és transformat en alcohol etílic; és així com el most esdevé vi (fermentació alcohòlica). De fet, al raïm premsat no cal afegir-li llevat perquè ja en té per ell mateix, per això els grans tenen aquest aspecte opac que els caracteritza (pruïna).
El nucli
Si ens esgarrapem una mà, les cèl·lules lesionades són substituïdes per altres cèl·lules semblants, de tal manera que la marca ni tan sols es veu. En néixer, fem una mitjana de 50 cm de llarg, i a tretze anys gairebé hem triplicat la nostra alçada. Si plantem una mongetera, al cap de poc temps tindrem una planta les llavors de la qual desenvoluparan altres mongeteres semblants a la primera. D’una gata neixen gatets, d’una gla una alzina, i dels nostres pares hem nascut nosaltres, que ens hi assemblem.
Tots aquests esdeveniments, i molts d’altres, tan habituals i quotidians, en realitat són causats per mecanismes extraordinàriament complexos i eficients, el desenvolupament dels quals és regulat pel nucli cel·lular.
Funcions i composició
El nucli acompleix un conjunt de funcions que es poden esquematitzar de la manera següent:
a) determina el fet que d’una cèl·lula concreta en neixin d’altres del mateix tipus: per exemple d’una cèl·lula de la pell altres cèl·lules epitelials, etc.;
b) regula l’activitat de la cèl·lula, en particular la formació de les proteïnes i d’altres compostos;
c) determina la diferenciació cel·lular, que va del zigot (que és l’òvul fecundat, o sigui la primera cèl·lula d’un nou organisme) a la formació d’un organisme complex amb milers de milions de cèl·lules. Són cèl·lules diferents segons la funció, i estan organitzades en teixits i òrgans especialitzats i coordinats entre ells. També controla, segons un pla ben determinat, el creixement, el desenvolupament i l’envelliment dels éssers vius.
El nucli és present en totes les cèl·lules eucariotes (excepte en els glòbuls vermells de la sang, que no tenen nucli i no es reprodueixen), i és ben visible al microscopi si es tenyeix amb els colorants adients. Hi ha cèl·lules que tenen més d’un nucli. Generalment és un corpuscle esferoïdal, delimitat per una membrana. Amb el microscopi electrònic s’hi poden distingir el nuclèol i la cromatina.
El nuclèol és un amàs més o menys arrodonit de grànuls i filaments amb abundant RNA, important molècula que intervé en la síntesi de les proteïnes (de les quals es parlarà a "La síntesi de les proteïnes").
La cromatina és formada per proteïnes i DNA. Quan la cèl·lula no s’està dividint es presenta en forma de filaments prims, constituïts per una altra molècula fonamental, el DNA, i per proteïnes. Durant la divisió cel·lular, els filaments, que tracen espirals, originen els cromosomes.
La cromatina i els cromosomes
La cromatina és constituïda, com ja s’ha dit, per dos components, aproximadament en la mateixa quantitat: proteïnes i DNA. Les proteïnes són força petites i s’anomenen histones. N’hi ha de cinc menes i són diferents per a cada espècie. El DNA és una llarga molècula doble, com veurem millor més endavant. La cromatina, al microscopi electrònic, sembla un collaret de perles: cada perla és un nucleosoma, constituït per un grup d’histones al voltant del qual es replega el DNA.
Durant la divisió cel·lular, cada filament de cromatina s’enrotlla en espiral estretament, s’escurça i augmenta de gruix. És així com es formen els cromosomes, uns corpuscles del nucli. Es poden veure fàcilment al microscopi òptic; n’hi ha prou d’agafar un fragment de l’extrem d’una arrel, per exemple la d’una ceba que hagi estat en remull alguns dies. L’arrel creix molt ràpidament i, per tant, té moltes cèl·lules que es divideixen. Es comprimeix el tros d’arrel delicadament entre dos portaobjectes i s’hi afegeix un colorant (per exemple carmí acètic). En mirar al microscopi, serà fàcil veure a l’interior dels cromosomes de moltes cèl·lules diferents moments de la divisió cel·lular (la mitosi).
Els cromosomes són formats per dos filaments, les cromàtides, cadascun dels quals conté una molècula de DNA. Les cromàtides s’uneixen en un punt on hi ha una estrangulació, el centròmer, que també és el punt on conflueixen les fibres del fus mitòtic durant la divisió cel·lular. A vegades hi pot haver estrangulacions secundàries.
Cada espècie viva té un nombre constant de cromosomes que és igual per a cada individu de la mateixa espècie. El conjunt dels cromosomes característics de cada individu constitueix el cariotip (de la paraula grega "cario", nucli).
Totes les cèl·lules del cos d’un organisme tenen idèntic cariotip. Cadascuna de les nostres cèl·lules somàtiques té 46 cromosomes, totes les cèl·lules somàtiques dels gossos en tenen 78, totes les dels gats en tenen 38, i així successivament. Les cèl·lules somàtiques sempre tenen un nombre parell de cromosomes (dotació cromosòmica diploide, simbolitzada per 2n). Els humans, i gairebé tots els animals, tenen una sèrie de parells de cromosomes iguals (autosomes) i un parell de cromosomes anomenats cromosomes sexuals o heterocromosomes que determinen el sexe de l’individu. Si els cromosomes sexuals són iguals (dos cromosomes en forma de X; per tant, XX) l’individu és femella; si són diferents (un cromosoma en forma de X i un altre en forma de Y; per tant, XY), l’individu és mascle. Així, els homes tenen 22 parells d’autosomes i un parell de cromosomes sexuals XY. Les dones tenen 22 parells d’autosomes i un parell de cromosomes sexuals XX.
Cada cromosoma és portador de determinats gens, és a dir, de les indicacions relatives als caràcters hereditaris de l’individu. Aquest tema es desenvolupa amb més profunditat a "Les lleis de la genètica".
El DNA
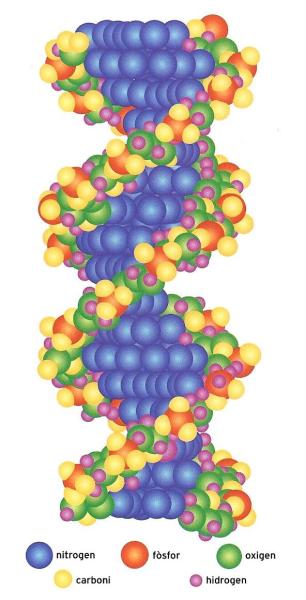
Representació simplificada d'un segment de molècula de DNA, l'element constitutiu dels cromosomes, que conté tota la informació genètica de l'organisme. Consisteix bàsicament en una doble cadena macromolecular composta de molècules del sucre desoxiribosa i de fosfats (les cintes helicoïdals més externes) i quatre tipus de bases nitrogenades (al cilindre central).
ECSA
La forma i el nombre dels cromosomes són, doncs, iguals per a tots els individus de la mateixa espècie. En l’interior dels cromosomes, però, el DNA de cada organisme és diferent del de tots els altres. No hi ha al món dos éssers amb un DNA igual. És precisament l’especificitat del DNA el que fa que cadascú de nosaltres sigui diferent de la resta. El DNA, portador dels caràcters hereditaris, és, doncs, igual en totes les nostres cèl·lules, però diferent del de qualsevol altre individu.
Quan una cèl·lula es reprodueix, transmet a totes dues cèl·lules filles una parella completa del seu DNA, és a dir, els transmet les instruccions que serveixen per a fer que la cèl·lula funcioni per ella mateixa i perquè funcioni tot l’organisme. Immediatament abans de la duplicació cel·lular (de la qual es parla en el paràgraf següent) cada cromosoma es duplica i forma així una còpia de la informació que conté. Cada cèl·lula filla rep un nombre igual de cromosomes i un patrimoni d’instruccions genètiques idèntic.
En les cèl·lules sexuals, en canvi, el nombre de cromosomes és la meitat del de les cèl·lules de la resta del cos, les cèl·lules somàtiques. En l’home els espermatozoides tenen 22 autosomes i un cromosoma X o un cromosoma Y, i en les dones els òvuls tenen 22 autosomes i un cromosoma X. De la fecundació d’un òvul per un espermatozoide amb cromosoma Y s’obté, per tant, un individu mascle, mentre que si l’espermatozoide conté el cromosoma X en sortirà una femella. La meitat dels 46 cromosomes de les cèl·lules dels fills, totes derivades del zigot, seran d’origen matern, i l’altra meitat d’origen patern (vegeu també "Les lleis de la genètica").
El DNA es duplica separant, com una cremallera que s’obre, les dues cadenes que el formen. Després, cada filament reprodueix un filament idèntic al que s’ha separat, ja que cada adenina s’uneix solament amb la timina, i cada guanina amb la citosina. El resultat és la formació de dues cadenes idèntiques a les anteriors; ara hi ha dues molècules amb el mateix DNA. El DNA fa dues funcions que són fonamentals per a tots els éssers vius. D’una banda, conté la informació necessària que permet la síntesi de totes les proteïnes d’un organisme i s’ocupa de fer-la arribar als ribosomes, on aquesta síntesi es produeix degudament descodificada. De l’altra, es duplica amb la finalitat que cada cèl·lula filla contingui la mateixa dotació genètica que la cèl·lula mare.
Com és el DNA
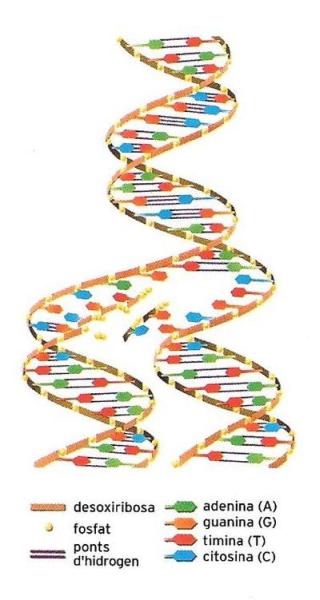
Esquema de l'estructura de la molècula de DNA, on es veu la disposició aparellada de les bases nitrogenades i la manera com l'hèlix s'obre per a permetre'n l'autoduplicació. L'estructura de la doble hèlix amb les bases dirigides a l'interior podria fer pensar que la molècula té una estructura rígida, i en canvi és a contrari: es tracta d'una molècula molt versàtil, que pot adoptar configuracions molt diferents, segons quina funció realitza i amb quina altra molècula es relaciona.
ECSA
L’àcid desoxiribonucleic (DNA) és una molècula molt gran, formada per moltes unitats elementals unides entre elles. Aquestes unitats s’anomenen nucleòtids.
Cada nucleòtid és compost per un sucre (la desoxiribosa), un grup fosfòric i una base. Les bases són de dues menes: les purines, d’anell doble, que són l’adenina i la guanina, i les pirimidines, d’anell simple, que són la citosina i la timina. Hi ha, doncs, 4 tipus de nucleòtids, un per a cada tipus de base. Un nucleòtid té com a base l’adenina, un altre la guanina, un altre la citosina i l’últim la timina. El grup fosfòric d’un nucleòtid s’uneix amb el sucre d’un altre, i forma una seqüència. El DNA és constituït per dues llargues cadenes de nucleòtids unides a través d’enllaços dèbils establerts entre les bases. Les bases, però, tan sols es poden unir d’una certa manera: la guanina amb la citosina, i la timina amb l’adenina. Es pot representar la molècula del DNA com una escala de cargol on els parells de bases serien els travessers, i la seqüència sucre-grup fosfòric, els muntants. Les dues cadenes de nucleòtids s’enrotllen sobre ells mateixes. Per tant, la seqüència vertical dels nucleòtids pot ser formada pràcticament per infinites combinacions, però els enllaços laterals entre les dues cadenes són forçosos: adenina (A) ≡ timina (T) i citosina (C) = guanina (G) (cada guionet indica un enllaç).
La seqüència dels nucleòtids determina la transmissió d’un caràcter o d’un altre, com una mena de "codi", que precisament s’anomena codi genètic. Simplificant es pot dir, per exemple, que els ulls d’una persona són blaus o marrons segons com siguin els nucleòtids de la zona del DNA que determina el color dels ulls. L’especificitat dels enllaços entre les bases, en canvi, dóna al DNA la possibilitat d’autocopiar-se (la còpia obtinguda és idèntica).
Vida i mort de les cèl·lules
Les cèl·lules neixen, creixen, formen diverses estructures, dupliquen els seus cromosomes i, finalment, es reprodueixen. El període que va del naixement a la duplicació s’anomena interfase. Les cèl·lules del nostre cos tenen cicles vitals diferents. El funcionament harmònic d’un organisme és garantit per l’alternança coordinada de reproducció i interfase de les seves cèl·lules. Algunes cèl·lules humanes i d’altres animals no es dupliquen. Els glòbuls vermells, per exemple, mancats de nucli, viuen una mitjana de 4 mesos i són renovats per les cèl·lules de la medul·la òssia, que en produeixen uns 2 milions i mig per segon. D’altres cèl·lules es poden reproduir intensament si l’organisme ho requereix. Per exemple, si a un individu només li quedés una tercera part del fetge, les seves cèl·lules hepàtiques es reproduirien fins a reconstruir un òrgan de mida normal.
Es pot calcular que en un ésser humà adult es produeixen 2 3 1012 divisions cel·lulars en 24 hores. En el temps que trigueu a llegir aquesta frase, uns 100 milions de les vostres cèl·lules s’han dividit (uns 25 milions per segon).
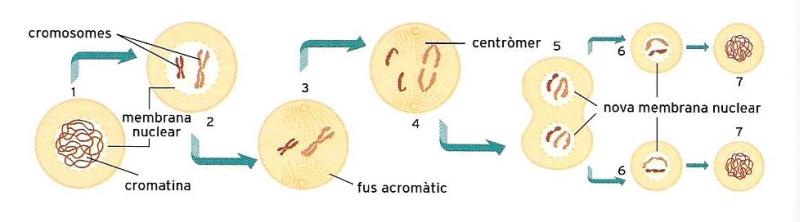
Esquema molt simplificat de la divisió cel·lular: (1) transformacions al nucli cel·lular i desaparició de la membrana nuclear, (2) duplicació dels cromosomes, (3) formació del fus acromàtic, (4) migració dels cromosomes aparellats als dos pols cel·lulars, (5) divisió del citoplasma i segmentació cel·lular, (6) formació de la membrana nuclear i (7) retorn dels cromosomes a l'estat de fibra de cromatina.
ECSA
Però les cèl·lules, per bé que es divideixin, no es conserven sempre joves. El seu envelliment és inevitable i irreversible, està vinculat a l’edat de l’organisme. En realitat no se sap gairebé res sobre les causes de l’envelliment, i de moment encara no s’ha aconseguit impedir-lo o retardar-lo. Quan una cèl·lula envelleix es transforma profundament, la membrana esdevé menys permeable, s’acumulen rebuigs del metabolisme i els orgànuls cel·lulars s’alteren.
Quan un ésser humà mor, moren unes 1014 cèl·lules (un 1 seguit de 14 zeros!, és a dir, 100 bilions); són les que formen el seu cos. N’hi ha que han viscut molt poc, perquè han substituït altres cèl·lules de les quals són filles. La pell humana, per exemple, és composta per cèl·lules de vida curta, és a dir, capaces de reproduir-se contínuament. En canvi, hi ha cèl·lules que viuen molt, tant com nosaltres. Es tracta sobretot de les cèl·lules nervioses, que envelleixen lentament i són capaces d’acomplir les seves funcions sense dividir-se mai. Hi ha una cèl·lula, però, que en certa manera és immortal. Es tracta de la cèl·lula sexual, que, quan origina un nou organisme, genera totes les seves cèl·lules, inclosos els seus gàmetes, els quals podran formar altres éssers vius i així successivament. Amb els protoctists passa el mateix, si fa no fa: poden morir, però no pas de vells (una cèl·lula vella dóna naixença a dues de joves o més).
Un cop les cèl·lules han assolit un volum, ja no poden continuar creixent sense arriscar-se a morir; aleshores es divideixen. Les cèl·lules somàtiques produeixen cèl·lules amb el seu mateix nombre de cromosomes; en aquest cas, es parla de mitosi. La meiosi, en canvi, és una reproducció cel·lular particular que produeix les cèl·lules sexuals, o gàmetes, amb la meitat dels cromosomes. La meiosi es descriu a "Les lleis de la genètica", perquè les lleis que regeixen la transmissió dels caràcters es basen en el comportament dels cromosomes durant la meiosi.
La mitosi té una durada variable, entre 40 minuts i una hora i mitja aproximadament, segons les cèl·lules. Els cromosomes arriben a la mitosi amb el DNA ja duplicat; cadascun és format per dues cromàtides, cadascuna de les quals conté una molècula de DNA. Un cop desaparegut l’embolcall nuclear els centríols i l’astrosfera s’allunyen mútuament i se situen un a cada pol de la cèl·lula, deixant entre ells un feix de fibres anomenat fus mitòtic, on es col·loquen els cromosomes durant la fase anomenada metafase. Aquest és el moment en què els cromosomes, que presenten les seves dues cromàtides en forma de X, són més fàcils de visualitzar. Després, els cinetòcors de cada cromosoma se separen i cadascun arrossega una cromàtide fins a un pol del fus, de manera que la meitat de les cromàtides emigra cap a un pol de la cèl·lula i l’altra meitat cap a l’altre.
Seguidament, mentre es reconstrueixen els dos nuclis nous als pols de la cèl·lula, també es produeix la divisió del citoplasma o citocinesi. Així es formen dues cèl·lules amb un patrimoni genètic idèntic al de la cèl·lula mare i aproximadament amb la meitat del seu citoplasma i els seus corpuscles. Tot i que aquest és el procés més corrent, de vegades es dóna la mitosi sense la citocinesi, i s’obtenen cèl·lules plurinucleades.
La síntesi de les proteïnes
Allò que som depèn en bona mesura de les proteïnes presents en el nostre cos. Els nostres cabells són arrissats o llisos segons com són les proteïnes que els formen; tan sols digerim la llet si al nostre estómac hi ha l’enzim específic, que és una proteïna; el fet d’agafar la varicel·la depèn solament de la presència o no a la nostra sang dels anticossos específics, és a dir, les proteïnes capaces de protegir-nos d’aquesta malaltia; que sapiguem una cosa depèn només de si al nostre cervell les proteïnes de determinades neurones han estat modificades per la informació en qüestió o no. El DNA determina les nostres característiques en la mesura que determina les nostres proteïnes. Per què es formen unes determinades proteïnes i no unes altres? Com es formen?
La composició de les proteïnes
Les proteïnes són cadenes constituïdes per 20 compostos orgànics anomenats aminoàcids. L’alfabet té 26 lletres que es poden encadenar per a formar paraules en un nombre pràcticament infinit de combinacions. Els 20 aminoàcids també es poden combinar de múltiples maneres. Però no totes les seqüències de lletres tenen significat o tenen el mateix significat; cal que es combinin en un ordre ben determinat.
Les lletres CSAO, ordenades d’aquesta manera, no signifiquen res; però si les disposem així: COSA, o així: OSCA, o així: SOCA, adquireixen significats ben concrets i diferents. De la mateixa manera, els aminoàcids s’han de disposar en una seqüència ben precisa per a formar una determinada proteïna (per a la composició de les proteïnes vegeu també "La gran família del carboni").
En les nostres cèl·lules hi ha un mecanisme que controla la formació de les proteïnes, mecanisme que es basa en un codi, el codi genètic, i en dues molècules: el DNA i l’RNA.
L'acid ribonucleic: RNA
El DNA, del qual ja hem parlat anteriorment, conté i transmet la informació genètica, però la molècula que forma materialment les proteïnes del citoplasma és l’RNA (àcid ribonucleic). Té la mateixa estructura que el DNA, amb algunes diferències: en comptes del sucre desoxiribosa, té un altre sucre de 5 àtoms de carboni, la ribosa, i en comptes de la timina té l’uracil. Els seus nucleòtids, doncs, poden contenir adenina, citosina, guanina i uracil. A més, mentre que el DNA és una hèlix doble, l’RNA és format per una sola cadena de nucleòtids. El DNA transcriu la seva informació a l’RNA, és a dir, forma un RNA que conté els mateixos nucleòtids (a banda de les diferències esmentades abans). De la mateixa manera que un filament de DNA en forma un altre de complementari, a partir d’un d’aquests filaments del DNA també s’obté l’RNA, que no és més que una còpia formada segons la norma de l’aparellament de bases: adenina (A) ≡ uracil (U) i citosina (C) = guanina (G) (com hem vist en l’insert "Com és el DNA"). L’RNA es forma en el nucli i després passa al citoplasma.
Hi ha tres tipus de RNA:
1) RNA missatger (RNAm). És un llarg filament de milers de nucleòtids, còpia del DNA.
2) RNA transferidor (RNAt). És una molècula petita, replegada sobre ella mateixa, capaç d’unir-se d’una banda a l’RNAm i, de l’altra, a un aminoàcid específic. Hi ha més de 20 tipus d’RNAt, almenys un per a cada aminoàcid.
3) RNA ribosòmic (RNAr). Forma, juntament amb unes proteïnes, els ribosomes i s’uneix, d’una banda, a l’RNAm i, de l’altra, a l’RNAt.
El codi genètic
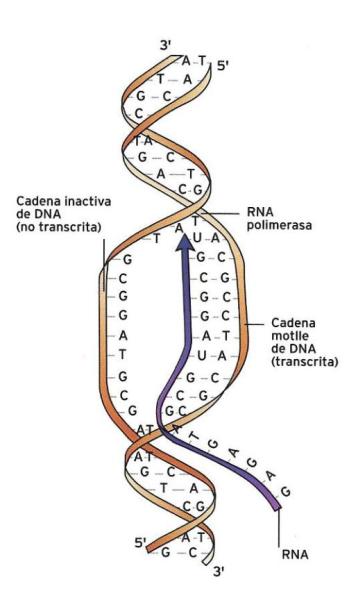
La transcripció del codi genètic és la primera etapa de l'expressió gènica mitjançant la qual es copia la informació del DNA en el llenguatge de l'RNA. Aquest procés consisteix en la síntesi d'una molècula d'RNA a partir dels seus corresponents precursors i amb la intervenció d'un enzim (l'RNA polimerasa) utilitzant com a motlle una de les dues cadenes del segment de DNA (gen) que ha de transcriure.
ECSA
Tal com ja hem vist, el DNA controla la seqüència dels aminoàcids de les proteïnes mitjançant l’RNA. Però l’RNA conté solament 4 elements variables, és a dir, 4 tipus de nucleòtids diferents. Com s’ho fa l’RNA, amb un "diccionari" de 4 lletres, per a reconèixer-ne 20, tantes com aminoàcids?
L’RNA és format per una cadena constituïda per quatre anelles diferents. Cada anella és un nucleòtid. Les proteïnes són també llargs compostos químics formats per una vintena de constituents, que són els aminoàcids. No es pot establir, doncs, una correspondència directa entre cada nucleòtid i cada aminoàcid, perquè els quatre nucleòtids existents només podrien reconèixer quatre aminoàcids i, com sabem, les proteïnes són combinacions de fins a vint aminoàcids. Aquest "sistema" tampoc no funcionaria si calgués dos nucleòtids per a reconèixer un aminoàcid; en aquest cas les combinacions només serien 16, és a dir, 42. La solució és que tres nucleòtids (que constitueixen un triplet) corresponen a un aminoàcid. Hi ha, doncs, 43, és a dir 64 combinacions possibles, anomenades codons, per a indicar els codis per als aminoàcids. Això significa que a un mateix aminoàcid li poden correspondre un, dos, tres, quatre o fins i tot sis codons, és a dir que hi ha "sinònims" que indiquen el mateix aminoàcid. L’aminoàcid triptòfan, per exemple, té com a codó UGG i, en canvi, l’alanina en té quatre; GCU, CGG, GCA, GCG (U = uracil, A = adenina, G = guanina, C = citosina). Dels 64 codons, solament 61 corresponen a aminoàcids. Els tres triplets restants (UAA, UAG, UGA) no corresponen a cap aminoàcid, sinó que indiquen el final del missatge, o sigui el punt on s’interromp la cadena, perquè la proteïna és completa.
La construcció de les proteïnes
Resumim breument tot el que hem dit fins ara: tots els éssers vius contenen DNA i RNA; la informació genètica del DNA es transcriu a l’RNA, que la tradueix en proteïnes; el codi de traducció es basa en la correspondència entre tres nucleòtids de l’RNA i un aminoàcid. A la pràctica, com s’esdevé la formació de les proteïnes en el citoplasma?
La síntesi proteica s’esdevé en els ribosomes, en els quals es col·loca l’RNA missatger com una cinta transportadora que conté la seqüència de codons corresponents a cada proteïna. Els aminoàcids són transportats per l’RNAm i col·locats en correspondència amb els seus codons. L’RNAt "captura" els aminoàcids del citoplasma i els transporta. Cada tipus d’RNAt en reconeix un. La molècula d’RNAt té en un extrem un codó complementari del de l’RNAm (anomenat anticodó), la qual cosa li permet reconèixer el triplet específic de l’aminoàcid de l’RNAm; a l’altre extrem de la molècula hi ha una zona que només s’uneix a un aminoàcid concret.
Els aminoàcids, doncs, es troben ordenats en l’RNA segons la seqüència determinada pel mateix RNAm i, per tant, pel DNA. En aquest moment els aminoàcids s’uneixen els uns als altres (enllaç peptídic) i formen una llarga cadena. Quan a l’RNAm, que es desplaça pel ribosoma, hi apareix el triplet que marca el final de la seqüència, aquesta se separa: la proteïna és acabada. En aquest moment, tant l’RNAm com els RNAt estan a punt per a formar-ne una altra d’idèntica a la primera. Així se sintetitzen moltes cadenes proteiques iguals, seguint el mateix ordre.
Un dels grans avenços de la biologia ha estat la comprensió d’aquest mecanisme complex i precís. Aquest procés té lloc en tots els organismes, cosa que demostra la unitat substancial de tots els organismes vius. En aquest moment quasi totes les cèl·lules del nostre cos estan treballant en la síntesi de noves proteïnes, i el mateix es pot dir de les cèl·lules del nostre gat, dels protozous que neden en l’aigua del gerro de flors i fins i tot de les cèl·lules de les flors.
Fins i tot els bacteris que ens provoquen el mal de coll tan desagradable sintetitzen les seves proteïnes d’una forma anàloga, i així és com viuen i proliferen. Els antibiòtics, que ens guareixen ràpidament, actuen precisament "sabotejant" aquest mecanisme. Són substàncies produïdes per diversos microorganismes (els bacteris, les floridures i els fongs) capaços d’interferir en la síntesi proteica dels bacteris, però no en la nostra. L’estreptomicina, per exemple, provoca errors en la lectura de l’RNAm dels bacteris, la penicil·lina impedeix la formació de les molècules de la seva membrana cel·lular, etc.
Cèl·lules diferents: de l'especialització cel·lular als teixits i els òrgans
Com és que les cèl·lules d’un organisme, malgrat tenir totes elles els mateixos cromosomes i els mateixos gens, són diferents? És una pregunta per a la qual encara no hi ha una resposta del tot satisfactòria. Es considera que no tots els gens són actius en el mateix moment. En el DNA hi hauria dos tipus de gens, els estructurals, que regulen la síntesi de les proteïnes, i els reguladors, que controlen quins gens estructurals han de ser actius i quins no. Quan una cèl·lula es diferencia, hi hauria tota una sèrie de gens que quedaria inactiva.
De totes maneres, en el nostre cos hi ha diversos tipus de cèl·lules organitzades en teixits. Un teixit és un conjunt de cèl·lules amb la mateixa funció. Es distingeixen 4 tipus de teixits: epitelial, muscular, nerviós i connectiu.
El teixit epitelial és una capa contínua de cèl·lules que revesteix el nostre cos, les seves cavitats i els seus canals. Tot allò que entra al nostre organisme, en surt o hi circula passa a través del teixit epitelial. Les seves cèl·lules, d’una o més capes, poden ser cúbiques o cilíndriques. Moltes (les cèl·lules glandulars) estan especialitzades en la producció de substàncies concretes com la saliva, la suor, la llet, les hormones, el moc i els enzims.
El teixit muscular és format per cèl·lules capaces de contreure’s, gràcies a la presència de certes proteïnes com l’actina i la miosina, que, en lliscar l’una sobre l’altra, escurcen la fibra muscular (vegeu també "Els mecanismes del moviment"). Hi ha dos tipus de músculs: els músculs llisos i els músculs esquelètics, segons si les molècules proteiques es disposen de manera regular o no, per constituir estries. Els músculs llisos, que no presenten estries, són els dels òrgans interns, com l’aparell digestiu o l’úter. És contreuen sense que hi intervingui la nostra voluntat, per això també reben el nom de músculs involuntaris. Els músculs esquelètics s’insereixen en els ossos i el seu moviment és controlat per la voluntat; per aquest motiu també s’anomenen músculs voluntaris. Són formats per milers de fibres, unides pel teixit connectiu. Cada fibra és una cèl·lula amb molts nuclis. Un múscul esquelètic específic és el que constitueix la paret del cor, la contracció del qual no depèn de la voluntat, sinó que és regulada per un mecanisme autònom.
El teixit nerviós és constituït per les neurones i les cèl·lules de la glia; aquestes darreres nodreixen i protegeixen les neurones, que són les autèntiques cèl·lules nervioses (vegeu també "La regulació de l'organisme"). Les neurones estan especialitzades en la recepció i la transmissió dels senyals del medi intern o extern, que envien en forma d’impulsos a altres neurones o a altres òrgans (els músculs, les glàndules, etc.). La neurona té un cos central (amb el nucli i els orgànuls citoplasmàtics), un conjunt de ramificacions anomenades dendrites, que reben els impulsos, i l’axó, que transmet aquests impulsos i els allunya del cos cel·lular.
Les neurones poden ser molt grans; l’axó d’una neurona motriu va des de la medul·la espinal fins als dits dels peus. Les neurones són de tres tipus: sensorials, motrius i de connexió. Les primeres reben les sensacions i les transmeten, com per exemple les que porten les sensacions visuals de l’ull al cervell. Les neurones motrius s’encarreguen de conduir l’impuls fins als músculs i les glàndules, i en determinen l’activitat. Les neurones de connexió relacionen les altres neurones entre elles.
La denominació de teixit connectiu inclou tots aquells teixits que, tot i ser molt diferents, tenen la funció d’unir, sostenir i protegir la resta de teixits. Dins aquest grup hi distingim el teixit ossi, el teixit cartilaginós, el teixit connectiu fibril·lar, el teixit elàstic, etc. La sang i la limfa també pertanyen a aquest grup heterogeni.
El teixit connectiu conté relativament poques cèl·lules, les quals no estan en contacte sinó que queden separades per una substància intercel·lular, anomenada substància fonamental, produïda per les mateixes cèl·lules. És constituïda per un material amorf de consistència viscosa i mancat d’estructura, i per fibres que poden ser elàstiques (per exemple les dels grans vasos sanguinis), col·làgenes (les de la pell, els tendons i els lligaments) o reticulars (formen entramats reticulars a l’interior dels òrgans, com per exemple el fetge).
La cèl·lula vegetal
Considerem una cèl·lula vegetal "tipus", la més genèrica i comuna, i comparem-la amb la cèl·lula animal descrita anteriorment. Descobrirem que totes dues tenen els mateixos components bàsics que fan possible que les dues cèl·lules, l’animal i la vegetal, desenvolupin les funcions que les fan semblants.
La cèl·lula vegetal, com l’animal, té una membrana cel·lular (anomenada plasmalemma), zona de límit i intercanvi amb el medi extern, que inclou un citoplasma en el qual es reconeixen els mateixos orgànuls: els mitocondris, per al desenvolupament de la respiració cel·lular; els ribosomes i el reticle endoplasmàtic, per a la síntesi de les substàncies que cada cèl·lula és capaç de fabricar; l’aparell de Golgi, per a l’acumulació i la secreció dels seus productes cel·lulars, i, finalment, el nucli, seu del material genètic que, en el moment de la divisió cel·lular, s’organitza en cromosomes la forma i el nombre dels quals són característics de cada espècie.
Però les cèl·lules vegetals tenen alguna cosa que les fa diferents... Tenen alguna cosa de més i alguna cosa de menys.
Hi reconeixem tota una família d’orgànuls citoplasmàtics exclusius, els plastidis: alguns (els cloroplasts) són especialitzats en el desenvolupament de la fotosíntesi clorofíl·lica; d’altres (els amiloplasts), en l’acumulació del midó, substància de reserva per excel·lència dels vegetals, i n’hi ha d’altres (els cromoplasts) que contenen pigments diferents.
A part d’aquests orgànuls especials, la cèl·lula vegetal difereix d’una manera clara de l’animal quan és observada globalment, fins i tot amb poc augment, al microscopi òptic. El seu contorn apareix com una línia marcada en comptes d’una línia evanescent com en la cèl·lula animal. Efectivament, té la membrana "reforçada" exteriorment per una paret cel·lular més o menys gruixuda (el gruix oscil·la aproximadament entre una micra, és a dir, una mil·lèsima de mil·límetre, i algunes desenes de micra), formada principalment de cel·lulosa (almenys en les plantes superiors). A més, no sembla que el citoplasma ompli tot l’espai disponible a l’interior de la paret cel·lular. Aquest espai en bona part és ocupat per cavitats plenes de líquid: els vacúols (a vegades, només n’hi ha un de molt gran).
Durant la divisió cel·lular també s’observen diferències. En la cèl·lula vegetal no hi ha centríols i no es forma la centrosfera característica de les cèl·lules animals. A més, la rigidesa de la paret, per bé que relativament prima, impedeix que el cos cel·lular s’estranguli per la meitat per a separar les dues cèl·lules filles. Un cop dividits els nuclis, el citoplasma no es comença a dividir amb els seus moviments actius, sinó que es forma un envà divisori, la làmina mitjana, com a paret comuna a les dues noves cèl·lules que separa. La làmina comença a formar-se inicialment a la part central de la cèl·lula i després es va estenent cap a la perifèria fins atènyer les parets laterals de la cèl·lula dividida. Seguidament, les dues cèl·lules construeixen sobre aquesta làmina noves capes de paret, i així esdevenen del tot independents l’una de l’altra.
Quin és, doncs, el motiu de la presència d’aquesta paret cel·lular, que gairebé sembla una cuirassa rígida i enutjosa? La seva funció principal és protegir contra una absorció excessiva d’aigua. En efecte, el medi líquid que envolta les cèl·lules vegetals té normalment una concentració més baixa de sals que els líquids cel·lulars interns, i sense una paret rígida i poc dilatable la cèl·lula tendiria a absorbir aigua; s’aniria inflant cada cop més fins a rebentar!
Hi ha un segon motiu que també justifica la presència de la paret en les cèl·lules vegetals. Com que els vegetals no tenen cap estructura comparable a l’esquelet dels animals, cal que hi hagi una estructura de suport per a impedir que les plantes es dobleguin a causa del pes, i és aquesta la funció de les parets cel·lulars. Això explica que les gegantines sequoies puguin dreçar-se cap al cel, que els enormes baobabs es mantinguin drets i també que les llargues i primes tiges del blat puguin sostenir el pes aparentment excessiu de l’espiga carregada de fruits. Per a un major aprofundiment d’aquests elements específics i distintius de les cèl·lules vegetals, us remetem a "Les plantes del nostre entorn".
La cèl·lula procariota i els bacteris
La cèl·lula eucariota no és compartida per tots els éssers vius. Hom anomena procariota la cèl·lula que té el material genètic en el citoplasma, normalment en una sola molècula circular de DNA, i que no té orgànuls cel·lulars, per contraposició a la cèl·lula eucariota, que té el material genètic (DNA) contingut en un nucli envoltat per una membrana i pot contenir diferents orgànuls cel·lulars en el seu citoplasma.
En la cèl·lula procariota típica poden observar-se unes estructures bàsiques, que són: la membrana cel·lular (o membrana plasmàtica), barrera que separa el medi intern cel·lular de l’ambient extern on viu; la paret cel·lular, coberta que envolta la membrana plasmàtica i que proporciona rigidesa a la cèl·lula, alhora que n’impedeix la lisi osmòtica, i el citoplasma, constituent fonamental d’una cèl·lula, delimitat per la membrana plasmàtica, i on hi ha gairebé tots els components cel·lulars. El citoplasma inclou: a) els ribosomes, petites partícules molt abundants (una sola cèl·lula procariota pot tenir fins 20 000 ribosomes) formades per àcid ribonucleic (RNA) i proteïnes, i és on es realitza la síntesi proteica; b) la regió nuclear (recordem que la cèl·lula procariota no té un nucli diferenciat), que és la zona on es concentra el material genètic o DNA, i que hom anomena "cromosoma" per analogia amb la cèl·lula eucariota; c) les inclusions, formades per materials de reserva, com ara alguns compostos de carboni, nitrogen, sofre o fòsfor, per gas (vacúols de gas), que els permet la flotació, per pigments (clorosomes) necessaris per a la fotosíntesi, partícules de ferro (magnetosomes), que permeten a la cèl·lula reaccionar davant de forces magnètiques, etc. Moltes cèl·lules procariotes poden moure’s, i ho solen fer mitjançant una estructura filiforme i contràctil, única o múltiple, anomenada flagel. Els flagels són una mena de motors que, amb el seu moviment rotatori, causen el desplaçament de la cèl·lula en un fluid. Pel que fa a la morfologia, les cèl·lules procariotes poden ser molt diferents les unes de les altres: rodones (cocs); en forma de bastonets cilíndrics (bacils), o corbats de manera senzilla (vibris), o en forma d’hèlix senzilla (espirils) o complicada (espiroquetes), o bé tenir formes irregulars (bacterioides). Poden presentar prolongacions cel·lulars amb diferents finalitats. Les fímbries, o pèls, poden ajudar la cèl·lula a adherir-se a una superfície. Un tipus especial de pèl sexual permet la unió de dos bacteris per a intercanviar material genètic, en el procés que hom anomena conjugació. Hi ha un grup especial de bacteris —les espiroquetes— que té uns flagels modificats, situats entre la paret i la membrana de la cèl·lula.
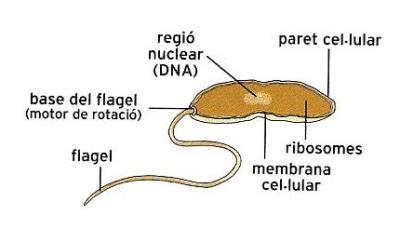
Organització cel·lular d'un bacteri tipus, que reuneix les característiques més generals d'aquests organismes. Sota la paret cel·lular rígida hi ha la membrana cel·lular, que tanca el citoplasma, i una zona central ocupada pel nucleoide, que no és envoltat de membrana. El flagel és un orgànul filamentós rígid i ondulat, fet de proteïna, que s'implanta a la membrana plasmàtica mitjançant el motor flagel·lar, que el fa rodar.
ECSA
La cèl·lula eucariota és normalment molt més gran que la procariota. Té un nucli (format per una membrana nuclear amb porus, cromosomes —amb DNA i proteïnes—, RNA, etc.), un citoplasma amb moltes estructures membranoses, uns ribosomes més grans que els de la cèl·lula procariota, mitocondris (orgànuls encarregats de la respiració) i cloroplasts (orgànuls amb clorofil·la de les cèl·lules que fan fotosíntesi, etc.). Però, a més de les diferències morfològiques, les cèl·lules procariotes i les eucariotes presenten també diferències en la seva composició química. La paret cel·lular característica de la majoria dels procariotes està formada principalment de peptidoglicà (anomenat també mureïna), compost que mai no es troba en els eucariotes.
La mida és un altre tret característic. La major part de les cèl·lules procariotes fan entre unes dècimes de micròmetre (µm; 1 µm= 10–6 m) i uns pocs µm. Les cèl·lules eucariotes són prop de deu vegades més grans. Unes dimensions petites permeten una velocitat de creixement i de multiplicació més grans gràcies a la relació existent entre la superfície cel·lular i el volum del seu citoplasma. La superfície determina la rapidesa amb què els nutrients poden entrar dins la cèl·lula, de manera que, com més gran és aquesta àrea, més ràpidament hi entren. I també, com més gran és el volum, més ràpidament s’utilitzen els nutrients per al manteniment, reparació, creixement i reproducció. Les cèl·lules petites tenen una relació superfície/volum més gran, és a dir, amb un mateix volum, tenen més superfície. Aquesta relació és per a un bacteri (cèl·lula procariota) típic vint vegades més gran que, per exemple, per a una cèl·lula humana (cèl·lula eucariota). Per tant, la cèl·lula procariota cobreix més fàcilment les seves necessitats nutricionals i creix més ràpidament. Per exemple, en condicions òptimes, el bacteri Escherichia coli es duplica cada vint minuts.
Els primers bacteris que ens vénen al pensament són potser els que ens provoquen certes malalties, com la tuberculosi, el tifus, el còlera, el tètanus, l’amigdalitis, etc., perquè són paràsits de les nostres cèl·lules. Però la majoria dels bacteris no són perjudicials; n’hi ha molts que són útils o indispensables per als altres éssers vius. Ocupen llocs essencials en les cadenes alimentàries, sovint permeten la vida d’altres organismes a l’interior dels quals viuen en simbiosi, o intervenen en el procés de fer comestibles substàncies que no en serien. El nostre intestí, per exemple, allotja bacteris que constitueixen la flora intestinal i que produeixen algunes substàncies que ens són útils (les vitamines). Diversos bacteris provoquen putrefaccions i fermentacions; determinen per exemple la transformació del vi en vinagre i la de la llet en iogurt, mantega o molts formatges. De tota manera, els més nombrosos són els que viuen al sòl, on descomponen les substàncies orgàniques i en permeten la reutilització per part de les plantes (i per tant dels animals que, directament o indirectament, es nodreixen de les plantes; vegeu també "Les plantes del nostre entorn"). Alguns bacteris produeixen substàncies que s’han revelat de gran utilitat en medicina. Es tracta de nombrosos antibiòtics que els humans han après a obtenir de manera controlada a partir dels bacteris, per a utilitzar-los en la cura de les malalties causades per altres espècies bacterianes.
Els bacteris poden tenir forma arrodonida (cocs), de bastonet (bacils), de coma (vibrions), d’espiral (espirils), d’hèlix, ramificada; a més, poden viure aïllats, en grup o en colònies.
Segons la seva necessitat d’oxigen, els bacteris se subdivideixen en anaerobis obligats i en aerobis. Els primers només poden viure en absència d’oxigen, i els segons solament si hi ha oxigen. A més, hi ha bacteris anaerobis facultatius (que també poden viure en presència d’oxigen) i bacteris microaeròfils (que viuen en ambients amb molt poc oxigen).
Els bacteris poden obtenir l’energia per a viure de moltes maneres diferents. Alguns bacteris són autòtrofs, és a dir, capaços d’obtenir la matèria orgànica que els és necessària per mitjà de processos de transformació de les substàncies minerals (alguns escindeixen compostos del sofre, altres del ferro, o del nitrogen) o per mitjà de processos de fotosíntesi parcialment diferents dels que permeten la fotosíntesi de les plantes (els bacteris no tenen clorofil·la, sinó certs pigments fotosensibles que s’anomenen bactericlorofil·les). D’altres bacteris, com els paràsits o els descomponedors, són heteròtrofs, perquè obtenen l’energia necessària per a viure a partir de la matèria orgànica produïda per altres organismes. Alguns són sapròfits, és a dir que es nodreixen de matèria orgànica morta, i d’altres són paràsits, obtenen aquestes substàncies a partir d’organismes vius.
Molts bacteris resisteixen les condicions ambientals adverses sota la forma d’espores. És el que es dóna, per exemple, en els bacteris del tètanus, malaltia que es pot contreure a través de les ferides infectades per la terra, els metalls rovellats, etc., que continguin precisament espores tetàniques. Les espores es formen a l’interior del citoplasma de la cèl·lula bacteriana, i després d’una fase de creixement són alliberades a l’exterior del bacteri, que es destrueix immediatament. De cada espora només s’origina un bacteri.
Els cianobacteris, també anomenats algues blaves, presenten un model cel·lular semblant. Viuen en l’aigua dolça, aïllats i, més freqüentment, en grups o units per llargues cadenes. Són més especialitzats que els bacteris, perquè són exclusivament autòtrofs: obtenen l’energia per a viure per mitjà de la fotosíntesi. En efecte, el seu citoplasma conté clorofil·la (la mateixa que les plantes, però no està organitzada en cloroplasts com passa en les cèl·lules vegetals).
Així els cianobacteris produeixen la matèria orgànica que els és necessària i l’oxigen. Molts cianobacteris són capaços de transformar el nitrogen de l’atmosfera en compostos orgànics (vegeu "El sistema viu"). La seva presència en els arrossars del sud-est asiàtic, per exemple, permet un cultiu continu d’arròs sense haver-hi d’afegir cap tipus de fertilitzant.
La filogènia molecular en la classificació dels éssers vius
Euriarqueotes.
ECSA
Les modernes tècniques de la biologia molecular permeten classificar els éssers vius a partir de les seves afinitats filogenètiques, mentre que les classificacions clàssiques anteriors es basaven principalment en relacions morfològiques i, especialment en els procariotes, també fisiològiques. Així doncs, els éssers vius s'havien classificat en cinc regnes, dels quals únicament un és procariòtic (regne moners). La filogènia molecular estableix que la vida cel·lular va evolucionar al llarg de tres llinatges principals, que hom ha denominat dominis. Hi ha dos dominis que són microbians, i comprenen exclusivament cèl·lules procariotes (domini Bacteria, també anomenats eubacteris, i domini Arquea, també anomenats arqueobacteris; el tercer és eucariòtic (Eucaria, també anomenats eucariotes). L'arrel de l'arbre universal filogenètic representa un moment del temps evolutiu on la vida que podia haver-hi a la Terra provenia d'un mateix avantpassat, a partir del qual l'evolució va prendre dues direccions, la que correspon al domini Bacteria i la dels Arquea-Eucaria. Aquests dos dominis se separen més endavant en dues línies diferents. Cal destacar que filogenèticament aquests dos dominis són més propers entre ells que amb el domini Bacteria. La filogènia ha permès confirmar algunes conclusions que s'havien fet mitjançant la taxonomia clàssica, com ara la major antiguitat dels arqueobacteris, que se separaren del tronc comú més a prop de l'arrel. Inclou per tant els organismes més primitius, mentre que el domini (Eucaria (on hi ha tots els protists, les plantes, els fongs i on trobem els animals) és ara el més allunyat de l'avantpassat comú i és el més evolucionat.
L'origen dels procariotes
L’aparició de les primeres formes de vida, que podrien considerar-se avantpassats directes de l’actual cèl·lula procariòtica, pot haver-se esdevingut fa uns 3 800 milions d’anys, quan la superfície de la Terra (que té una edat d’uns 4 550 milions d’anys) es devia refredar fins a una temperatura que permetés la presència d’aigua líquida, substància indispensable per a l’aparició i el manteniment de la vida. En roques de fa 3 500 milions d’anys s’han detectat microfòssils que contenen bacteris força evolucionats, amb una estructura molt semblant a la d’alguns microorganismes actuals. D'altra banda, en les roques més antigues que es coneixen, la formació d’Isua, a Grenlàndia, amb una edat d’uns 3 850 milions d’anys, no s’hi han detectat cèl·lules però sí algunes molècules que podrien ésser indicis d’activitat prebiològica.
L’aparició de formes vives molt senzilles es creu que va poder tenir lloc moltes vegades mitjançant associacions que haurien format protocèl·lules constituïdes potser tan sols per una membrana lipídica que contingués un àcid nucleic (RNA) primitiu amb capacitat de síntesi i algunes proteïnes. Es desconeix el pas que va dur de la previda a la vida, i que va fer possible que un conjunt de substàncies químiques reunides de manera aleatòria adquirissin les característiques que defineixen un sistema viu, és a dir, la capacitat d’automanteniment i d’autoreplicació, constituint allò que hom anomena un sistema autopoètic. La cèl·lula és el primer sistema autopoètic. Ara bé, si perquè aparegués aquesta primera cèl·lula calien unes condicions adequades, també van ser necessàries per al seu manteniment. És possible, per tant, que la vida hagi sorgit més d’un cop a la Terra. I potser també en algun altre planeta del sistema solar, per exemple a Mart. La vida que coneixem és l’única de la qual tenim constància, però no és pas l’única possible.
Les primeres cèl·lules devien nodrir-se de les molècules orgàniques del medi per a mantenir la seva estructura, obtenir-ne energia i multiplicar-se. En augmentar la quantitat de cèl·lules, van haver de desenvolupar mecanismes metabòlics que permetessin el reciclatge, de manera que els productes que resultaven del metabolisme d’uns organismes servissin d’aliment a d’altres. Altrament, les molècules orgàniques acumulades a la Terra s’haurien esgotat en uns 200 o 300 milions d’anys.
Els organismes poden classificar-se segons el seu tipus de metabolisme, tenint en compte la font d’energia (fototròfics o quimiotròfics), la font de poder reductor (litotròfics o organotròfics), la font de carboni (autotròfics o heterotròfics) i, finalment, segons la substància acceptora d’electrons (respiradors aeròbics, respiradors anaeròbics o fermentadors). Els procariotes (és a dir, els organismes que tenen cèl·lules procariòtiques, com ara els bacteris) són l’únic grup d’éssers vius que presenta tots els tipus possibles de metabolisme.
El cultiu dels bacteris
La bacteriologia és la ciència que estudia els bacteris. És una branca de la microbiologia que amplia el seu camp a tots els éssers vius microscòpics (els anomenats "microbis"), des dels virus fins als moners i els protoctists.
Un dels capítols més interessants de la tècnica bacteriològica és el del cultiu dels bacteris, que consisteix en el trasplantament d’aquests microorganismes, procedents dels medis naturals més variats, en medis nutritius adequats anomenats brous de cultiu, que permeten que sobrevisquin i es multipliquin. Aquest cultiu és molt important perquè per ell mateix (és a dir, només amb els cultius de laboratori) fa possible l’estudi sistemàtic i aprofundit de la biologia de les diverses espècies.
Els brous de cultiu poden ser més o menys complexos i en l’actualitat n’existeixen moltíssims, cadascun dels quals és particularment indicat segons les circumstàncies i l’espècie que es vol cultivar. Poden ser líquids o sòlids; els que habitualment s’utilitzen en els laboratoris de bacteriologia són el brou, la gelatina i l’agar-agar.
El brou és un medi líquid que s’obté fent bullir carn magra sense greix ni tendons i afegint-hi sal i peptona (una substància proteica). Al brou es desenvolupen quasi totes les espècies bacterianes; el seu creixement queda demostrat per l’enterboliment del líquid o per l’aparició d’una capa superficial prima. La gelatina és una substància proteica extreta dels ossos, dels cartílags i de la pell d’animals mitjançant l’ebullició en aigua; un cop preparada es conserva a 20°C per evitar-ne la liqüefacció. Els gèrmens que es desenvolupen en la gelatina poden créixer en superfície i formar colònies arrodonides més o menys extenses, o bé restar al fons i originar imatges característiques, diferents entre una i altra espècie, com la d’avet invertit (Bacillus anthracis) o d’embut (Staphylococcus aureus). L’agar-agar és un cultiu sòlid, constituït per una substància mucilaginosa extreta d’algues a la qual s’afegeix peptona i clorur de sodi. Permet el cultiu de la majoria dels microorganismes. Els bacteris hi creixen formant colònies superficials, de forma i grandària diferents, de vegades de colors. Aquestes característiques poden ser utilitzades per a un primer reconeixement de l’espècie bacteriana aïllada.
Els brous de cultiu, un cop preparats, han de ser esterilitzats, és a dir, tractats de manera que no hi quedi vida; així, quan el brou s’utilitza, tan sols hi creixen els microbis que s’han introduït expressament. L’esterilització s’aconsegueix per mitjà de la temperatura o bé per filtració.
És interessant destacar que les cèl·lules bacterianes que són trasplantades a un brou de cultiu no comencen a multiplicar-se immediatament després del trasplantament, sinó al cap d’un cert temps, que s’anomena període de latència. Durant aquest període, les cèl·lules no es multipliquen, però hi ha una intensa activitat metabòlica principalment representada per la síntesi de tots els enzims necessaris per a la utilització dels substrats continguts en el brou. Arriba un moment que aquest treball preparatori cessa, i les cèl·lules comencen a multiplicar-se, primer lentament i després cada vegada més de pressa (fase d’acceleració). Aquesta fase dura més o menys, però en un cert moment es comença a notar un alentiment (fase de desacceleració o d’alentiment) que porta la població bacteriana a una situació d’equilibri, en la qual el nombre de bacteris per unitat de volum roman constant, i les cèl·lules que moren són substituïdes per les que neixen. El cultiu entra en una fase estacionària de durada variable però que en comporta inevitablement un debilitament, causat d’una banda per l’exhauriment dels substrats nutritius necessaris per a la multiplicació bacteriana i, de l’altra, per l’acumulació de substàncies tòxiques que acaben obstaculitzant la reproducció cel·lular.
Els virus
Morfologia d'un virus madur totalment format o virió: un nucli central constituït per DNA o RNA i una coberta proteica anomenada càpside, que en els més complexos és feta de nombroses subunitats o capsòmers. Malgrat la seva simplicitat estructural, les seves formes i dimensions són molt variables. Aquesta simplicitat i la manca de metabolisme propi els situa a la frontera entre les coses vives i les inertes.
ECSA
Els virus poden ser considerats microorganismes, però no formen part de cap regne, perquè són incapaços de viure independents. Són paràsits obligats de les cèl·lules, les quals en molts casos eliminen, i causen malalties a les plantes i als animals; alguns fins i tot són paràsits de bacteris, és a dir, d’altres microorganismes. Són malalties víriques, per exemple, la grip, la varicel·la i el xarampió. Els virus són formats per un àcid nucleic (DNA o RNA) embolcallat per una coberta proteica. En un virus mai no coexisteixen els dos àcids nucleics, i aquest fet condiciona els virus fent-los incapaços de sintetitzar proteïnes i, per tant, de ser autònoms.
La coberta proteica fa que cada tipus de virus pugui penetrar en un únic tipus de cèl·lules i no en d’altres. Els virus del refredat, per exemple, solament envaeixen les cèl·lules de la mucosa del nostre aparell respiratori. Un cop han penetrat en la cèl·lula, l’àcid nucleic del virus, que conté la informació genètica per a la síntesi dels enzims necessaris per a la reproducció (replicació) del microorganisme, utilitza les estructures de la cèl·lula hoste per a reproduir-se milers de vegades. Els bacteriòfags, virus paràsits de bacteris, només injecten DNA en l’hoste; la cèl·lula bacteriana el duplica diverses vegades, i forma nous bacteriòfags que s’alliberen quan es destrueix el bacteri.