Una panoràmica dels vegetals
El "món vegetal" inclou una enorme varietat d’organismes, sovint molt diferents entre ells: des dels microorganismes formats per una cèl·lula, o unes quantes, fins als arbres majestuosos de cent metres d’alçada o més; de les fràgils herbes mogudes pel vent al tronc robust i capaç de resistir la fúria dels elements; de la planta de cicle vital curt a la que viu durant segles... Aquestes imatges són tan sols un exemple de la gran varietat del món vegetal.
Abans de començar a examinar com són i com viuen els organismes vegetals és important intentar fer-ne una classificació general. Aquesta classificació ha de posar de manifest, més enllà de les diferències quant a l’aspecte extern, les semblances o diferències bàsiques pel que fa a l’estructura i les funcions vitals dels diversos grups vegetals.
Així, una primera divisió general proposaria dos grans grups: els tal·lòfits i els cormòfits (noms derivats de tal·lus i corm respectivament), perquè podem descriure l’estructura del cos (el soma) d’un organisme vegetal a partir d’aquests dos models bàsics. El tal·lus no presenta una diferenciació del cos en diversos òrgans (o, si aquesta diferenciació existeix, és extremament limitada). En el corm, en canvi, el soma es divideix en tres òrgans principals: l’arrel, la tija i la fulla, cadascun dels quals té funcions i característiques diferents.
Els tal·lòfits
El tal·lus és l’estructura típica de tots els "vegetals inferiors", que amb raó podem considerar que van aparèixer al planeta en èpoques anteriors. Es tracta en primer lloc de les algues d’aigües dolces i salades, tant unicel·lulars com pluricel·lulars.
Per a les cèl·lules l’aigua és un bon medi líquid on alimentar-se, respirar i excretar sense necessitat de tenir parts del cos especialitzades en l’absorció, el transport de materials d’una part a l’altra de l’organisme, etc. I tampoc no és indispensable que altres parts es transformin per a ancorar-se al fons (per bé que a vegades passi), o per a sostenir el cos perquè es mantingui erecte. En efecte, el vegetal pot mantenir la posició vertical gràcies a la flotació aconseguida per un equilibri exacte entre les parts que s’estenen (com si fossin les fulles) i les parts primes o amb veritables òrgans de flotació. El pas de l’aigua a la terra ferma no significa la pèrdua de l’estructura tal·losa, però sí la seva limitació. Hi ha plantes terrestres amb tal·lus, però totes són petites (d’uns centímetres d’alçada o poc més) i viuen en llocs molt humits, de manera que no han de penetrar en el sòl per a trobar aigua; quan tenen estructures semblants a arrels (que s’anomenen "rizoides"), les fan servir sobretot per a fixar-se al sòl.
Són tal·lòfits terrestres els briòfits, més coneguts com a molses i hepàtiques. Es tracta de plantetes constituïdes per una petita tija erecta o reptant, dotada de rizoides encarregats de la fixació al sòl i de l’absorció, i de fulletes disposades en espiral en tres sèries. Viuen en colònies grans que de vegades ocupen extensions notables. També tenen estructura de tal·lòfit els fongs (que en realitat, per les diferències que presenten respecte als altres organismes del regne vegetal —principalment la incapacitat de realitzar la fotosíntesi clorofíl·lica, per la qual cosa no són organismes autòtrofs, sinó heteròtrofs—, generalment s’agrupen en un regne a part; vegeu "Els fongs") i els líquens, organismes derivats de la simbiosi entre algues i fongs.
Les algues El grup principal de tal·lòfits aquàtics és el que anomenem genèricament algues. En primer lloc hem de parlar de les cianobacteris (també anomenades cianofícies o algues blaves). Són organismes unicel·lulars o que formen grups gelatinosos o filaments gairebé sempre no ramificats, que viuen preferentment en les aigües dolces o sobre superfícies humides. Si bé es diferencien de la resta de les algues pel fet de ser organismes procariòtics (sense nucli diferenciat ni orgànuls cel·lulars), són tradicionalment estudiades pels algòlegs perquè sovint viuen barrejades amb les altres algues. Presenten un color verd blavós o verd maragda, degut a la presència d’un pigment blau, la ficocianina. La paret apareix sovint coberta per una capa gelificada. Les cianofícies són els organismes fotosintetitzadors més antics de la biosfera, en la qual van predominar durant més de tres mil milions d’anys, fins que van aparèixer els primers organismes eucariòtics. La resta de les algues presenten nucli i orgànuls cel·lulars, entre els quals destaquen els cloroplasts, que poden tenir organització i coloració diversa segons els pigments que acompanyen la clorofil·la. Un primer grup d’algues són unicel·lulars o formen estructures senzilles, i viuen sobretot al plàncton marí, com ara els dinòfits, organismes principalment unicel·lulars que presenten flagels, o preferentment en aigües dolces, com els crisòfits o algues daurades, més abundants a les aigües pures, o els euglenòfits, unicel·lulars amb un sol flagel i una taca vermella vers la base d’aquest, que tenen els cloroplasts de color verd fulla i viuen en aigües riques en matèria orgànica. Un destacat grup d’algues, principalment unicel·lulars i sense flagels en la fase vegetativa, és el de les diatomees, amb moltes espècies en les aigües dolces i una gran importància en el plàncton marí. Els plastidis tenen color verd brunenc i sovint acumulen gotes d’oli com a substàncies de reserva, però el caràcter principal és el frústul: un embolcall d’aspecte de vidre (format per sílice), integrat per dues meitats molt ornamentades, que encaixen com les dues parts d’una capseta.
Les algues brunes, anomenades així pel color verd brunenc (fins a negrós), causat sobretot per un pigment brunenc afegit a la clorofil·la, viuen principalment a les costes de les mars fredes i són notables perquè arriben a presentar una organització molt complexa, fins i tot amb cèl·lules conductores, i una biomassa i una producció prou grans per a poder ser explotades en grans quantitats per a extreure’n els alginats (substàncies emulgents), o per a emprar-les com a adob i fins i tot en l’alimentació humana. Al litoral de la Mediterrània podem trobar, a poca profunditat, comunitats de l’alga bruna Cystoseira, mentre que a les costes atlàntiques hi ha els Fucus, que queden emergits durant la marea baixa.
El grup d’algues marines més important, sobretot a les mars més o menys càlides, és el de les algues vermelles, caracteritzades per la presència d’un pigment vermell (ficoeritrina) i un de blau (ficocianina) que, afegits a la clorofil·la, els donen colors variats que van des del rosat, passant pel vermell viu, fins al violaci fosc. Les espècies del gènere Gelidium subministren el preuat agar i les de Gigartina i Chondrus proporcionen un altre valuós gelificant, la carraguenina. Però les més fàcils de veure a les nostres costes, just al nivell de l’aigua, són les Rissoella.
Hem deixat per al final les algues verdes, molt semblants per diversos caràcters (cèl·lules reproductores, cloroplasts de color verd fulla) als briòfits i les plantes superiors, que degueren evolucionar, a partir d’algun dels seus grups més complexos, per a iniciar l’aventura de la colonització del medi terrestre. Tot i això, les algues verdes són un grup fonamentalment d’aigua dolça, només secundàriament adaptat per a viure a la mar o sobre superfícies humides, que inclou des de representants unicel·lulars, de creixement ràpid, com les clamidomones, amb dos flagels apicals, o les clorel·les, sense flagels, que viuen a les aigües riques en sals minerals i que, juntament amb altres espècies, són responsables del color verd de les basses. Moltes són filamentoses, com les espirogires, i formen part del llot o verdet dels rius o aigües embassades. Les aigües marines afectades per l’activitat humana (ports, platges freqüentades) mostren poblacions de clorofícies foliàcies, com l’enciam de mar (Ulva) i les Enteromorpha.
Els briòfits Es fa difícil saber com degueren ser les primeres plantes que colonitzaren el medi terrestre. De tota manera podem dir que, en l’actualitat, els organismes més simples adaptats al medi terrestre són els briòfits. Aquests organismes, que comprenen les molses i les hepàtiques, no tenen un sistema conductor ben desenvolupat i perden fàcilment l’aigua que contenen. Per aquesta raó la majoria de les espècies viuen en indrets humits i poc assolellats. Amb tot, algunes espècies han après a resistir deshidratacions periòdiques i a recuperar l’activitat quan tornen a rebre aigua, de la pluja o de la rosada.
El cicle vital dels briòfits és un bon exemple de cicle de les plantes. Una de les fases, el gametòfit (el productor de gàmetes), és la que està més desenvolupada, i d’ella depèn la realització de la fotosíntesi. Pot tenir una estructura bàsica formada per una mena de tija (caulidi) amb una mena de fulles (fil·lidis), generalment formades per una sola capa de cèl·lules, com passa a les molses i a les hepàtiques folioses, o bé una estructura tal·losa, més semblant a una mena d’alga plana i allargada, com passa a les hepàtiques tal·loses. En qualsevol cas, hi ha uns filaments de fixació que col·laboren en la captació d’aigua per capil·laritat, però, en general, l’aigua i les sals minerals s’absorbeixen per tota la superfície.
La generació formadora d’espores (l’esporòfit) és una planteta sense fil·lidis i sense clorofil·la, o amb ben poca, que viu paràsita sobre el gametòfit, d’on treu els aliments fotosintetitzats. Sovint és de vida curta. A l’extrem superior hi ha un esporangi, allargat a les molses o esfèric a les hepàtiques, on es produeixen les espores. Les molses són molt diverses, però es caracteritzen, en general, per la presència de fil·lidis tot al voltant del caulidi. N’hi ha de petites, que formen coixinets sobre les pedres (Grimmia), o gespes de poca alçada, com les Tortula de les parets o la molsa d’estrelles (Pleurochaete), totes dues resistents a l’assecament, mentre que en indrets més humits n’hi ha de més grans, ramificades i reptants, que formen catifes, com les molses de pessebre (Hypnum i Pseudoscleropodium) dels nostres alzinars ombrívols, o presenten l’aspecte de coixins gruixuts, com la molsa de pisos (Hylocomium) dels nostres boscos subalpins.
Les hepàtiques folioses posseeixen fil·lidis a banda i banda del caulidi i acostumen a reptar sobre superfícies ombrívoles, de troncs (Frullania) o talussos (Lophocolea). Les hepàtiques tal·loses formen llengües verdes ramificades dicotòmicament sobre terra humida i ombrívola. Algunes tenen la superfície llisa i una estructura senzilla (Pellia), però les més característiques presenten la superfície reticulada i amb porus (Conocephalum). Entre les espores hi ha uns filaments que, en assecar-se, es disparen com una molla, fent esclatar l’esporangi i dispersant les espores.
Les algues
Les algues constitueixen un ampli i important grup de plantes adaptades a la vida en medis aquàtics o com a mínim humits. El seu cos s’organitza com un tal·lus. Aquesta estructura de tal·lus, l’activitat fotosintetitzadora i la vida aquàtica són els únics trets que tenen en comú totes les algues. Les altres característiques, en canvi, poden variar moltíssim. Hi ha, per exemple, algues unicel·lulars que fan una centèsima de mil·límetre, i algues pluricel·lulars molt grosses i amb una organització que recorda la de les plantes amb corm. Un exemple són les gegantines algues brunes que viuen al fons dels oceans i que, al llarg de les costes californianes, formen veritables boscos sota l’aigua, constituïts per individus que poden arribar als 100 m d’alçada o més. També són diferents els pigments a partir dels quals capturen la llum solar per a realitzar la fotosíntesi. Sovint, el color de la clorofil·la queda emmascarat pels pigments que hi ha al seu costat, els quals donen a l’alga coloracions que van del groc al bru; es parla d’algues verdes, brunes, vermelles i daurades. Aquesta coloració diferent de les algues respon a exigències molt concretes. En efecte, mentre que per a les algues que viuen a la superfície una coloració verda o entre groga i verda resulta favorable per a absorbir la llum útil per a la fotosíntesi, a les algues que viuen a més profunditat (on la llum és filtrada per les capes d’aigua superiors) els cal pigments vermells o bruns que absorbeixin precisament les radiacions que aconsegueixen penetrar a més profunditat.
En les classificacions modernes les algues unicel·lulars són incloses, juntament amb els protozous, en el regne dels protoctists, i constitueixen un component important del plàncton d’aigües dolces i marines. N’hi ha moltes que tenen un o dos flagels per moure’s; d’altres, com les diatomees, posseeixen un delicat involucre silícic que conté el cos cel·lular. Antigament els cianobacteris o algues blaves també s’incloïen dins les algues unicel·lulars, però la microscòpia electrònica ha revelat que són en realitat cèl·lules procariotes. Per tant, actualment s’inclouen en el regne dels moners (vegeu també "Vida i evolució"). Totes les algues tenen una importància extraordinària per als altres éssers vius perquè la seva activitat fotosintètica és bàsica per a la vida dels ambients aquàtics. En efecte, són les algues les encarregades de l’oxigenació de les aigües i de la producció contínua de substàncies orgàniques. Si no hi hagués algues, que amb la fotosíntesi alliberen oxigen contínuament, ben aviat els oceans, els llacs i els rius esdevindrien inhabitables per a qualsevol mena d’animal aquàtic. Igualment, les algues són la base de la cadena alimentària de tots els ambients aquàtics. El plàncton vegetal (el fitoplàncton) serveix d’aliment als peixos i al plàncton animal que, al seu torn, és aprofitat pels peixos. Es calcula que la quantitat de carboni convertit en forma orgànica per les algues en un any pot arribar als 100 o 150 mil milions de t, una quantitat 8 vegades més gran que tot el carboni produït pel conjunt de les plantes terrestres.
Ara bé, l’altra cara de la moneda és que les algues també poden ser perjudicials. En efecte, poden causar danys a conseqüència dels processos de putrefacció que segueixen els períodes de gran desenvolupament de determinades espècies (de resultes de la contaminació de les aigües), i també per l’existència d’espècies d’algues molt tòxiques. Una alga molt tòxica és l’alga unicel·lular marina Gonyaulax catenella, que pot créixer molt, fins al punt de crear grans masses vermelloses surant per la mar; aleshores es poden produir enverinaments per ingestió de musclos o altres mol·luscs que s’hagin alimentat d’aquesta alga.
També cal remarcar la importància de les algues per a l’economia humana. Sobretot al llarg de les costes japoneses es conreen nombroses algues bentòniques (dels fons marins) que s’utilitzen per a l’alimentació humana; d’altres s’empren com a farratge per als animals, o com a fertilitzants. Hi ha diverses espècies d’algues vermelles i brunes a partir de les quals s’obtenen productes emprats des de fa temps per la medicina, la indústria alimentària i els laboratoris científics (l’agar-agar, els alginats, etc.).
Els cormòfits
L’estructura amb corm s’interpreta com una resposta evolutiva dels organismes vegetals a les difícils condicions de vida de la terra ferma. En efecte, trobem una estructura tal·losa més perfeccionada, que ja s’acosta a la dels cormòfits, en plantetes de terra ferma (les molses). Però com que la seva estructura no és sinó una simple "imitació" d’un corm, aquestes plantetes no deixen de ser plantes minúscules.
Com ja s’ha dit anteriorment, l’estructura de les plantes amb corm (cormòfits) es divideix en tres parts principals: l’arrel, la tija i la fulla.
La raó per la qual les plantes terrestres —a diferència de les aquàtiques— han desenvolupat aquesta diferenciació rau en el fet que tenen el cos repartit, per dir-ho d’alguna manera, entre dos medis molt distints, el sòl i l’aire, dels quals obtenen una part de les substàncies nutritives que els són necessàries.
Les plantes amb corm més antigues són els pteridòfits, que inclouen les falgueres, els equisets i els licopodis. Aquestes plantes ja tenen arrels, tiges i fulles, però es reprodueixen per espores com els altres grups vegetals "inferiors"; no tenen ni flors ni fruits ni llavors. Per tant, juntament amb els altres vegetals inferiors, s’anomenen criptògames (que en grec significa "de noces amagades"), però es distingeixen com a criptògames vasculars, perquè ja tenen veritables teixits, concretament els teixits destinats al transport de l’aigua i les substàncies que hi ha dissoltes, des de l’arrel fins a les branques i viceversa (els vasos conductors, d’aquí ve l’atribut "vascular").
Llevat d’algunes rares excepcions (les falgueres arbòries), els pteridòfits no superen el metre d’alçada. De fet, aquestes plantes no tenen la capacitat de reforçar les arrels i les tiges amb un creixement secundari tal com passa en els grups més evolucionats i, per tant, no poden tenir una estructura gaire gran.
La resta, nombrosíssima, de plantes s’agrupa en una sola categoria: els espermatòfits, és a dir, les plantes amb llavor o, segons la denominació de Linné, les fanerògames (del grec "de noces evidents", per contraposició a les criptògames), anomenades així perquè els òrgans reproductors són ben visibles. És precisament a partir dels òrgans femenins que es formarà la llavor.
Els espermatòfits es divideixen en dos grans grups: les gimnospermes i les angiospermes. En les primeres, les més nombroses són representades per les coníferes: els pins, els avets, les pícees, els xiprers, etc. la llavor no és dins un fruit, tot al contrari del que passa en les segones, totes les plantes amb flor.
Les angiospermes són les darreres plantes que van aparèixer a la Terra, i també les que tenen una estructura i unes funcions més complexes. Inclouen un nombre enorme d’espècies (més de 250 000), la majoria terrestres per bé que també n’hi ha d’aquàtiques. Són arbres, matolls, enfiladisses i herbes d’aspecte molt diferent, com també és diferent l’aspecte de les seves flors i els seus fruits.
Les angiospermes se subdivideixen en dos grups: les monocotiledònies i les dicotiledònies, el nom de les quals deriva d’una característica distintiva de la llavor. En les dicotiledònies, per exemple la mongetera, la llavor presenta dues "fulles" especials (els cotilèdons), que proporcionen l’aliment a l’embrió. En les monocotiledònies (per exemple, el blat) hi ha un sol cotilèdon. De tota manera, hi ha moltes més diferències entre aquests dos grups de plantes.
En els capítols següents estudiarem l’estructura de les diverses parts que constitueixen el cos dels cormòfits. Quant als òrgans reproductors, descriurem concretament les angiospermes, que presenten òrgans, com les flors i els fruits, que els són exclusius.
La cèl·lula vegetal
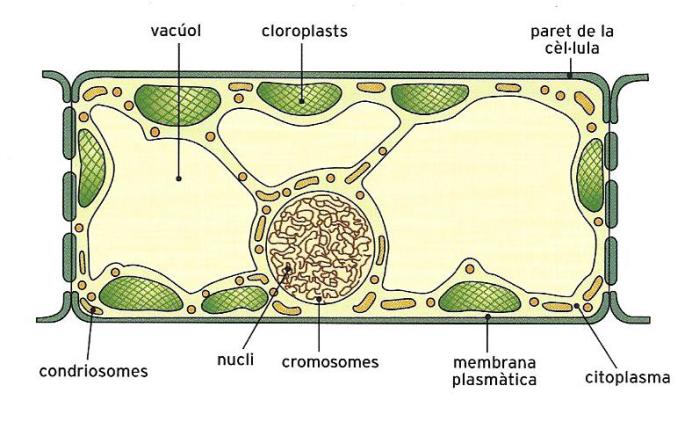
Esquema de la cèl·lula vegetal tipus, amb la paret cel·lular i un gran nombre de vacúols i plastidis de diferents menes que la diferencien de la cèl·lula animal.
ECSA
Els vegetals, com els animals, són constituïts per cèl·lules eucariotes. Les cèl·lules vegetals, però, per bé que tenen moltes coses en comú amb les animals, en difereixen en alguns aspectes importants (per a un estudi sobre la cèl·lula en general remetem a "La cèl·lula"). Així, presenten uns orgànuls i unes estructures que no tenen les cèl·lules animals: la paret cel·lular, els plastidis i el vacúol. Vegem ara en què consisteixen aquestes "especialitzacions" de les cèl·lules vegetals, i de quines maneres creixen i es desenvolupen.
La paret cel·lular
La cèl·lula vegetal és embolcallada per una mena de recobriment protector molt resistent però alhora elàstic (almenys, en les fases juvenils de la cèl·lula): la paret cel·lular. Un cop desenvolupada del tot, la paret cel·lular és constituïda per tres capes que, començant per l’exterior, són: la làmina mitjana, la paret primària i la paret secundària. La làmina mitjana és formada per un polisacàrid, és a dir, per un glúcid o un sucre complex, anomenat pectina. La paret primària es compon de filaments de cel·lulosa (el polisacàrid més abundant de les estructures vegetals) inclosos en una substància gelatinosa barreja d’altres polisacàrids i proteïnes. La paret secundària té els mateixos components que la paret primària, però amb més abundància de cel·lulosa.
Aquests components, que són comuns a totes les cèl·lules vegetals, es combinen amb altres components propis de cada teixit de la planta, segons les diferents funcions, i donen lloc a les anomenades "modificacions secundàries" de la paret cel·lular. Les més importants són: la lignificació, la suberització i la cutinització. La lignificació consisteix en la deposició de lignina, un polímer (en els teixits més durs constitueix més del 40% de la substància seca total) que proporciona rigidesa i resistència mecànica a la paret i que és típic, per bé que no exclusiu, del tronc. La suberització i la cutinització consisteixen, en canvi, en la deposició de suberina i cutina respectivament. Són substàncies impermeabilitzants que donen a la paret una gran resistència enfront dels agents químics.
La suberització sol afectar la totalitat de la paret i és típica de les cèl·lules del còrtex de la tija. El procés de suberització del còrtex, diferent en les diverses plantes, és molt més pronunciat en l’alzina surera, ja que la seva escorça és llevada periòdicament i aprofitada per a diferents aplicacions; una de les més comunes i més conegudes és la confecció de taps de suro. La cutinització, en canvi, normalment es limita a una sola cara de la cèl·lula, l’externa. És típica del teixit epidèrmic de les parts joves de la tija, les fulles, els fruits, els òrgans florals, etcètera.
Els plastidis
Són una família d’orgànuls cel·lulars que s’originen en els protoplastidis, petites vesícules indiferenciades presents en les cèl·lules joves dels meristemes, és a dir, els teixits encarregats del creixement de la planta. Posteriorment els protoplastidis s’engrandeixen i es diferencien, i adopten un aspecte i unes funcions específiques segons el teixit al qual pertanyen. Originen tres menes de plastidis: els cloroplasts, els cromoplasts i els amiloplasts.
Els cloroplasts es localitzen en òrgans i teixits verds (les cèl·lules foliars i les capes superficials verdes de les tiges joves), on la planta durà a terme la fotosíntesi clorofíl·lica. La seva estructura és complexa, i es caracteritza per tenir les membranes apilades les unes sobre les altres. Aquestes membranes són les que contenen la clorofil·la, organitzada en estructures altament complicades de les quals també formen part altres pigments (per exemple els carotenoides) i diverses substàncies necessàries per a la realització de la fotosíntesi.
Es consideren cromoplasts els cloroplasts degradats en els quals ha desaparegut la clorofil·la mentre que es conserven els altres pigments que donen a la cèl·lula, i per tant al teixit i a l’òrgan, la coloració vermella o groga. Aquests processos tenen lloc per exemple en molts fruits (els pebrots, els tomàquets, les llimones, les taronges, els plàtans, etc.), en les llavors (les granes del blat de moro) i en els pètals de les flors que són verds quan són immadures i després esdevenen vermells o grocs. Aquest mateix procés és el responsable de la variació tardoral del color de les fulles (esgrogueïment o envermelliment, segons l’espècie). En aquest cas, la degradació de la clorofil·la precedeix la degradació dels carotenoides, amb la qual cosa deixa d’haver-hi predomini del pigment verd sobre els altres pigments, que encara són presents (vegeu "Els colors de les plantes").
Els amiloplasts, també anomenats leucoplasts, són el magatzem on es conserva la substància nutritiva de reserva més important de la planta, el midó (del nom llatí del qual prenen la denominació aquests orgànuls). Els amiloplasts es troben en tots els teixits de la planta no exposats a la llum i en particular en els òrgans subterranis (les arrels, els bulbs, els tuber-cles i els rizomes), però també en els teixits profunds de la tija (la medul·la), les llavors, etc. En definitiva, allà on calgui acumular reserves.
El vacúol
Les cèl·lules vegetals adultes, observades al microscopi, presenten un aspecte totalment diferent de les cèl·lules animals perquè la major part del seu volum és ocupada pel vacúol en comptes del citoplasma, que es redueix a una pel·lícula fina adossada a la paret. El vacúol és una cavitat delimitada per una membrana i plena de líquid cel·lular, solució aquosa on es dissolen nombroses substàncies orgàniques i inorgàniques. Les funcions del vacúol són diverses. El seu paper fonamental és l’absorció de l’aigua segons els mecanismes osmòtics. Fa de "magatzem" de les substàncies que la planta es veu obligada a absorbir en excés. En una paraula, contribueix a regular la vida de la cèl·lula i la de tot l’organisme vegetal.
Creixement i desenvolupament de les cèl·lules
Per a descriure el creixement i el desenvolupament d’una cèl·lula vegetal convé començar per les cèl·lules anomenades "meristemàtiques", que es troben a l’àpex de les tiges i de les arrels, és a dir, a les parts més joves i més tendres d’aquests òrgans. Amb la seva divisió continuada, les cèl·lules meristemàtiques s’encarreguen d’augmentar el nombre de cèl·lules de l’individu, el fan créixer. Tots els òrgans vegetals capaços d’un creixement continuat tenen una zona de cèl·lules meristemàtiques. Aquestes cèl·lules són petites respecte de la mida mitjana de les altres i el seu aspecte és molt diferent del de la cèl·lula vegetal típica.
El creixement és conseqüència de la divisió cel·lular (la mitosi: vegeu "La cèl·lula"). Al final d’una mitosi, la cèl·lula meristemàtica s’ha dividit en dues cèl·lules filles la meitat de grosses que la cèl·lula mare, cadascuna de les quals creix fins a assolir les mateixes dimensions d’aquesta i després es divideix. Aquesta manera de créixer s’anomena creixement per divisió. S’hi poden distingir dues fases: la divisió pròpiament dita (augmenta el nombre de cèl·lules sense que variï la massa de matèria viva) i l’augment que equipara les dimensions de les filles a les de la mare.
Imaginem-nos que observem un grup de cèl·lules meristemàtiques, per exemple les de l’àpex d’una arrel. Aquestes cèl·lules es divideixen amb freqüència però, això no obstant, el seu nombre roman pràcticament constant al llarg del temps. Això es deu al fet que un cert nombre de cèl·lules abandona contínuament l’estat meristemàtic, per desenvolupar-se i transformar-se en cèl·lules adultes, incapaces de dividir-se. El pas d’una cèl·lula meristemàtica a una cèl·lula adulta implica 4 fets bàsics:
1) un gran augment de les dimensions, anomenat creixement per elongació (o elongació cel·lular);
2) un augment dels trets diferencials que distingeixen la cèl·lula vegetal de l’animal;
3) l’especialització d’una funció determinada;
4) la pèrdua de la capacitat de dividir-se, que tan sols es pot recuperar en casos excepcionals.
Mentre s’allarguen, les cèl·lules vegetals pateixen tota una sèrie de transformacions que es poden resumir en una sola paraula: diferenciació. Aquest terme indica la diversificació de les cèl·lules seguint moltes vies de desenvolupament divergents. De fet, a partir de les cèl·lules meristemàtiques —totes elles semblants— es desenvolupen molts tipus de cèl·lules adultes a les quals corresponen moltes funcions diferents. Segons l’"ofici" que tingui la cèl·lula un cop adulta, es desenvoluparà més una part o una altra durant la diferenciació. Per exemple, en una cèl·lula destinada a la fotosíntesi es desenvoluparan bàsicament els cloroplasts, mentre que la paret romandrà prima. En canvi, una cèl·lula destinada a servir de suport engruixirà la paret mentre que el citoplasma i els orgànuls es desenvoluparan poc o fins i tot desapareixeran; en aquest darrer cas, la cèl·lula morirà. En efecte, no és gens rar que la diferenciació comporti la mort de la cèl·lula: llavors, la cèl·lula assoleix la plena funcionalitat un cop morta. L’única part que aquestes cèl·lules conserven és la paret, per això la seva funció sempre està vinculada a aquesta part de la cèl·lula. Les típiques cèl·lules que tan sols funcionen un cop mortes són els vasos, que recorden tubs buits a través dels quals passa la saba.
La nutrició i el metabolisme de les plantes
Les plantes no són éssers estàtics, i no es poden descriure ni comprendre plenament si tan sols se’n coneix l’estructura. Com tots els organismes vius, acompleixen funcions vitals. L’estudi de les funcions vitals de les plantes és l’objectiu d’una disciplina de la botànica que rep el nom de fisiologia vegetal.
La fisiologia de les plantes és molt diferent de la dels animals, per bé que algunes grans funcions metabòliques són comunes a tots dos regnes. Les plantes, per exemple, no tenen aparell de recepció i conducció dels estímuls comparable al sistema nerviós dels animals i, per tant, no desenvolupen aquestes funcions. Es pot dir el mateix del sistema muscular, perquè, a diferència dels animals, són fixades al sòl (o a altres tipus de substrat) o com a màxim fluctuen sobre les aigües. Tanmateix, això no significa que no captin certs estímuls procedents del món exterior i que no tinguin la capacitat d’interactuar-hi, ni tampoc que siguin incapaces de fer moviments. És tan sols que fan aquestes funcions d’una manera completament diferent a la dels animals. En les plantes fins i tot són presents els fenòmens de regulació hormonal. És més, les hormones vegetals o fitohormones intervenen en les funcions més importants de la planta —l’auxina, per exemple, regula el creixement per elongació— i en la seva coordinació. Però en el món vegetal no hi ha estructures especialitzades en la producció i la secreció d’hormones com les glàndules endocrines dels animals. En canvi, hi ha una funció fonamental que tots els organismes han de satisfer per a sobreviure: la nutrició.
Els mecanismes de la nutrició
Des del punt de vista fisicoquímic, un ésser viu es pot considerar com un sistema que degrada i elimina de manera constant les seves parts constitutives (és a dir, les molècules biològiques que el formen). És per això que aquestes parts han de ser renovades contínuament. Els organismes s’encarreguen de substituir-les amb l’assimilació d’altres substàncies de l’exterior, que després, per mitjà de determinades seqüències de reaccions bioquímiques, esdevenen semblants a les molècules característiques de cada espècie: és el procés de la nutrició.
Segons el tipus d’organisme, aquest procés es desenvolupa seguint modalitats extremament variades, tant pel que fa a la naturalesa química de les substàncies nutritives (o nutrients) que s’absorbeixen, com a la manera d’incorporar-les i reelaborar-les. Els animals, per a aconseguir el recanvi de les molècules biològiques de què són constituïts, han d’absorbir altres molècules biològiques alimentant-se d’éssers vius, animals o vegetals. Per això són organismes heteròtrofs. En les plantes no és així, almenys pel que fa a la majoria. En efecte, són organismes autòtrofs: es nodreixen de substàncies no biològiques que absorbeixen de l’aire o del sòl, substàncies que reelaboren i recombinen per a obtenir substàncies biològiques noves.
La capacitat de convertir en viu allò que no ho és, de transformar en orgànic el que és inorgànic, és exclusiva de les plantes i s’anomena organificació. Són resultat d’aquest procés un nombre reduït de substàncies biològiques bàsiques que posteriorment són reelaborades i combinades per a formar la gran varietat de molècules que constitueixen fins i tot l’organisme més simple i primitiu.
Les plantes, doncs, són l’únic intermediari indispensable entre dos mons completament diferents: l’orgànic i l’inorgànic. A més, són el factor indispensable per a mantenir la vida animal, i per tant també la humana. Solament a través del procés d’organificació és possible renovar la substància orgànica que es degrada contínuament. Per tant, si l’organificació s’aturés, al cap de poc temps també desapareixeria la vida a la Terra. Així doncs, els animals sense les plantes no podrien sobreviure, mentre que, en general, les plantes sí que podrien viure sense els animals.
De què (i com) es nodreixen les plantes
Tots sabem que si ens oblidem de regar les plantes d’interior es panseixen i moren; i passa el mateix si deixem una planta massa temps allunyada de la llum del sol. Per a viure, doncs, les plantes necessiten llum i aigua. També sabem que les plantes, almenys les cultivades, s’han d’adobar de tant en tant. Però ningú no es preocupa d’anar a adobar les plantes d’un bosc, un prat, un bruguerar, etcètera.
Com es pot interpretar això? Segons les observacions anteriors, podríem afirmar que les plantes viuen gràcies a la llum i l’aigua, i que un adob natural o sintètic, per bé que no és estrictament necessari, les fa créixer millor.
Però aquesta afirmació, tot i que en certa manera és certa, no és suficient per a donar-nos una idea completa i, sobretot, lògica, de com es nodreixen les plantes. Cal fer una anàlisi científica més aprofundida, i aplicar les tècniques de l’anàlisi química. Aquesta anàlisi ens revela que els elements químics que constitueixen les plantes són els mateixos que constitueixen els animals (amb l’única excepció del sodi, que sembla que no té cap importància especial per als vegetals).
Que la constitució química sigui comuna entre els vegetals i animals és perfectament lògic, o més ben dit necessari, perquè els segons sempre són fets de matèria d’origen vegetal que han assimilat directament (és el cas dels herbívors) o indirectament (és el cas dels carnívors) a través d’altres animals que, al seu torn, s’han alimentat de vegetals. Les plantes, en canvi, es nodreixen de les substàncies minerals que troben en el medi on viuen i que després transformen en matèria orgànica.
Però, què s’entén per "hàbitat" de les plantes? Quines substàncies minerals assimilen exactament? A quines transformacions les sotmeten? Quins són els productes finals d’aquests processos de transformació?
En les plantes terrestres, el que s’anomena corm, l’estructura, es reparteix entre dos medis molt diferents: el sòl i l’atmosfera. Així, la tija i les fulles viuen en un ambient gasós, i les arrels en un medi d’estructura complexa, no solament sòlid, sinó també líquid i en part gasós. Les plantes obtenen les substàncies nutritives necessàries per a sobreviure tant de l’atmosfera com del sòl.
Les plantes i l’atmosfera. L’aire és una mescla gasosa en la qual el 78% és nitrogen, el 21% oxigen i l’1% restant inclou alguns altres gasos, com el diòxid de carboni, que en constitueix aproximadament el 0,03%. El nitrogen, el component més abundant, és un gas químicament inert. Des del punt de vista biològic, aquest element és de gran importància ja que intervé en la composició de les proteïnes i els àcids nucleics, però per a les plantes no representa normalment una font de subministrament (vegeu "El sistema viu").
L’oxigen és utilitzat directament per les plantes. Les cèl·lules vegetals, com les animals, respiren, o sigui, cremen les substàncies energètiques com els sucres i els greixos per obtenir l’energia necessària per als seus processos vitals. Però no pot ser considerat un veritable nutrient, encara que sigui indispensable per a la utilització de les substàncies nutritives. Entre les substàncies que constitueixen l’atmosfera, la que té una importància fonamental per a les plantes és el diòxid de carboni (CO2). Representa la font d’aprovisionament de l’element biològic més important: el carboni.
L’assimilació del CO2 per part de les plantes i la seva transformació en sucres d’alt contingut energètic gràcies a l’energia proporcionada per la llum solar constitueixen les etapes més importants d’aquest procés anomenat fotosíntesi clorofíl·lica (que estudiarem de manera més aprofundida en els apartats següents), tan fonamental perquè hi hagi vida a la Terra.
Les plantes i el sòl. Per a entendre algunes de les característiques bàsiques del sòl el podem comparar a un objecte familiar com pot ser una esponja. L’estructura del sòl no és compacta, sinó porosa, de manera que absorbeix l’aigua pels porus i la transmet a la superfície de les partícules que el constitueixen.
L’estructura d’un sòl, en essència, és una mescla de partícules sòlides de formes irregulars i dimensions diferents unides entre elles. Però la part sòlida que ocupen aquestes partícules no representa tot el volum disponible, perquè entre elles deixen espais de formes i dimensions diferents. Cada partícula sòlida és embolcallada per un vel d’aigua que omple completament els espais més petits, mentre que els més grans són plens de bombolles d’aire en contacte amb el vel aquós. El sòl, per tant, té una estructura complexa en la qual coexisteixen parts sòlides, líquides i gasoses estretament interconnectades les unes amb les altres. En créixer, les arrels de les plantes s’insinuen entre les partícules del sòl, on s’adhereixen.
En l’aigua hi ha dissoltes les sals minerals necessàries per a la nutrició de la planta. L’aire contingut en el sòl fa que les arrels puguin absorbir l’oxigen necessari per a la respiració.
Fins ara hem parlat de "sòl" en general, com si a tot arreu fos exactament igual. Però en realitat hi ha molts tipus de sòl, que, si bé tots responen a l’esbós anterior, tenen molts aspectes diferents, com la composició química i, molt important, la dimensió mitjana de les partícules sòlides. De fet, la classificació dels sòls es basa en aquest paràmetre i d’ell en depenen moltes de les propietats que són importants per a la vida de les plantes: la capacitat de retenció d’aigua, la permeabilitat, el contingut d’aigua i aire, etcètera.
La fotosíntesi clorofíl·lica
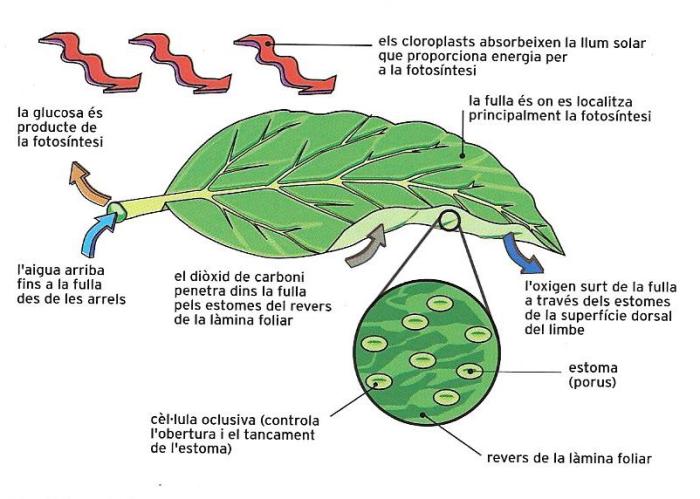
Les fulles són òrgans verds que tenen com a tasca principal realitzar la funció clorofíl·lica, és a dir, produir els composts orgànics que després són repartits per tota la planta, però també alliberar oxigen, que en part és reutilitzat en la respiració, i perdre vapor d'aigua a través dels estomes.
ECSA
La fotosíntesi clorofíl·lica és el procés a través del qual la planta assimila i organifica el carboni (és a dir, transforma la molècula inorgànica en molècula orgànica).
La fotosíntesi es basa en la transformació de dues substàncies inorgàniques d’estructura simple i de baix contingut energètic, el diòxid de carboni i l’aigua, en una substància orgànica complexa i d’alt contingut energètic, la glucosa. L’equació química de la fotosíntesi:
6CO2 + 6H2O → C6H12O6 + 6O2
ens diu que hi ha 6 molècules de diòxid de carboni (CO2) i 6 d’aigua (H2O) que es combinen per a formar 1 molècula de glucosa (que és un sucre, o glúcid, monosacàrid) i 6 molècules d’oxigen.
Tot i que l’oxigen és molt important per als organismes vius (les plantes incloses), des del punt de vista de la fotosíntesi es pot considerar un subproducte, mentre que el producte primari és la glucosa. En realitat, la glucosa no és l’única substància orgànica que es produeix al llarg del procés fotosintetitzador, ja que se n’originen moltes més (les proteïnes, els greixos i altres sucres) mitjançant processos de síntesi molt complicats. Però, com que és el producte més típic i abundant de la fotosíntesi, en tractar del procés de la fotosíntesi normalment s’hi al·ludeix.
No tota la glucosa produïda per la planta es "crema" immediatament per a satisfer-ne les necessitats metabòliques: una part es transforma en altres substàncies orgàniques que l’organisme vegetal utilitza de manera diferent, com la cel·lulosa i el midó. Concretament el midó s’emmagatzema en els cloroplasts durant el dia (midó primari) i durant la nit s’escindeix en sucres més simples que es transporten a zones concretes de la planta, on tornaran a adquirir la forma de midó (midó secundari) i actuaran com a substància nutrient de reserva, que s’utilitzarà en diversos moments del cicle vegetatiu.
Si observem l’equació general de la respiració cel·lular (vegeu "La cèl·lula"):
C6H12O6 + 6O2 → 6CO2 + 6H2O
ens adonem fàcilment que és exactament la inversa de l’equació de la fotosíntesi. De fet, es pot considerar que és una respiració inversa (o, viceversa, que la respiració és una fotosíntesi invertida): les substàncies que actuen de reactius de l’una són els productes de l’altra. La respiració cel·lular és un procés que allibera energia, la continguda en els compostos orgànics que es cremen. La fotosíntesi, al contrari, és un procés que acumula energia en quantitats que equivalen a la diferència entre la quantitat continguda en els reactius i la continguda en els productes.
Per tant, una manera més completa d’escriure les equacions de la fotosíntesi i de la respiració és la següent (on E és l’energia):
6CO2 + 6H2O + E → C6H12O6 + 6O2
fotosíntesi
C6H12O6 + 6O2 → 6CO2 + 6H2O + E
respiració
La font natural de l’energia utilitzada pel procés fotosintetitzador és la radiació lumínica del sol. En realitat, la llum no és sinó una forma d’energia i és gràcies a aquesta energia, distribuïda i difosa per tota la superfície de la Terra, que les plantes aconsegueixen realitzar la fotosíntesi.
Fins aquí, podem resumir tot el que hem dit anteriorment amb una afirmació important: l’hàbitat de la planta, i més exactament el de la seva part aèria (la tija i les fulles), inclou tots els factors necessaris per a la fotosíntesi: el diòxid de carboni contingut en l’aire atmosfèric, l’energia lumínica de la radiació solar i, l’últim reactiu necessari per a la fotosíntesi, l’aigua absorbida del sòl. L’aigua constitueix el dissolvent en què són dissoltes totes les substàncies cel·lulars i on hi ha "immersos", per dir-ho d’alguna manera, tots els òrgans cel·lulars, i la seva disponibilitat no representa, per tant, cap problema, entre altres coses perquè l’aigua que consumeix la fotosíntesi és tan sols una part petita de l’aigua total de la planta.
De quina manera interactuen aquests tres components, el diòxid de carboni, l’aigua i la llum, per a formar glucosa? En primer lloc cal explicar que la fotosíntesi es realitza en dues fases:
1) Una primera fase anomenada fase lumínica, perquè requereix la intervenció directa de la llum. En aquesta etapa la clorofil·la, pigment contingut en els cloroplasts de les cèl·lules vegetals, capta l’energia de la llum del sol i la transforma en energia química que contenen determinades molècules cel·lulars (NADPH i ATP). Aquestes molècules són una mena de "moneda energètica" directament utilitzable en els processos metabòlics que requereixen energia.
2) Una segona fase anomenada fase fosca, perquè, a diferència de la primera, no requereix llum. En aquesta etapa l’energia continguda en les molècules que hem anomenat "monedes energètiques" (NADPH i ATP) s’empra per a obtenir la glucosa (substància orgànica) a partir del diòxid de carboni i l’aigua (substàncies inorgàniques).
La fase lumínica de la fotosíntesi
La clau de tot el procés fotosintetitzador rau en última instància en la capacitat de la clorofil·la d’absorbir l’energia lumínica. Aquesta absorció provoca una modificació de l’estat energètic de la molècula de la clorofil·la, que aleshores és en condicions de poder cedir electrons (vegeu "La funció de la clorofil·la en el procés de la fotosíntesi"). En la primera reacció de la fase lumínica de la fotosíntesi un electró en estat excitat abandona una molècula de clorofil·la i passa a una altra molècula, que anomenem "acceptor primari" d’electrons. Aquesta reacció, que en condicions normals mai no tindria lloc, es realitza gràcies a l’empenta energètica proporcionada per la llum del sol, que permet a l’electró excitat, gràcies a l’energia suplementària emmagatzemada, trencar l’enllaç que l’uneix als nuclis de la clorofil·la i unir-se a una molècula (l’acceptor) que en condicions normals presenta una feble atracció electrònica.
Aquest procés es pot considerar del tot anàleg al procés pel qual s’aconsegueix fer arribar al cim d’un turonet un roc que inicialment era als peus d’aquest turó. Evidentment, això no és possible d’una manera espontània, però si li proporcionem prou energia (aixecant-lo amb un buldòzer o amb els braços), el roc podrà arribar al cim, encara que hagi de vèncer la força de la gravetat.
Un cop adquirit l’electró de la clorofil·la, l’acceptor primari d’electrons passa, al seu torn, a un estat excitat causat per l’energia que li ha transferit l’electró. Però una molècula excitada també és una molècula inestable. Com que en la natura totes les substàncies tendeixen a assolir l’estat de més estabilitat, o bé de mínima energia, l’acceptor primari també cedirà l’electró amb la seva energia a una altra molècula. El pas de l’electró de l’acceptor primari al "primer transportador" (una molècula anomenada ferredoxina) no és, per dir-ho d’alguna manera, un pas "ascendent". En aquest cas, l’electró passa d’una substància (l’acceptor) que el reté amb una certa força a la ferredoxina, que, per la seva estructura química, l’atreu amb una força més gran.
Però la ferredoxina no és l’objectiu final, és tan sols una anella de la cadena de molècules que "transporten" l’electró. Després de passar a través de la cadena dels transportadors d’electrons, cadascun dels quals adquireix l’electró del transportador precedent per cedir-lo al següent, l’electró és finalment cedit a una molècula cel·lular especial, anomenada NADP. Un cop rebut l’electró, les molècules d’NADP es comporten com totes les molècules que adquireixen electrons, és a dir, pateixen una reacció reductora que les transforma en molècules d’NADPH. El NADPH és un dels nucleòtids reduïts que proporcionen energia a les cèl·lules. La fase lumínica de la fotosíntesi produeix, doncs, un dels combustibles fonamentals dels organismes vius, l’NADPH.
Tot i això, no és l’únic "combustible" cel·lular produït per la fotosíntesi, que és capaç de generar-ne un altre: l’ATP. A "La cèl·lula" es descriu la importància de l’ATP pel que fa a l’acumulació i el transport d’energia, de la qual se serveixen les cèl·lules de tots els organismes. Ara bé, la cadena dels transportadors d’electrons del sistema fotosintetitzador inclou alguns passos entre els transportadors que comporten una caiguda d’energia suficient per a permetre la síntesi d’ATP, ADP i fosfat inorgànic. Aquesta síntesi s’anomena fotofosforilació.
El fet que les principals molècules que acumulen i intercanvien energia siguin les mateixes en organismes tan diferents com les plantes i els animals és una prova notable de la unitat constitutiva fonamental i de l’origen comú de tots els éssers vius.
Ara bé, se’ns pot ocórrer una observació. Durant la fase lumínica de la fotosíntesi hem vist que les molècules de clorofil·la perden electrons que, a través de la cadena de transportadors, atenyen el nucleòtid NADP i el transformen en NADPH.
Llavors, les molècules de clorofil·la, com es recarreguen d’electrons quan els en falten? Evidentment, hi ha d’haver en les cèl·lules una primera matèria que funcioni com a reserva d’electrons per a la fotosíntesi. Aquesta primera matèria no és cap molècula orgànica de fórmula complexa, sinó la més difusa i coneguda de les substàncies inorgàniques: l’aigua. Les molècules de clorofil·la, quan són abandonades per un electró en estat d’excitació i es troben, per tant, mancades d’electrons, exerceixen prou força per a atreure els electrons que uneixen els àtoms d’hidrogen i els d’oxigen de la molècula d’aigua. Això comporta, inevitablement, la ruptura d’aquests enllaços i la formació d’electrons (e), protons (H+) lliures i àtoms d’oxigen (O);
H2O → 2e- + 2H+ + O
Els àtoms d’oxigen, com que no poden quedar aïllats, de seguida es combinen entre ells per originar molècules d’oxigen diatòmic (O2) que ràpidament es difonen fora del cloroplast. L’oxigen produït per la fotosíntesi, essencial per a la vida a la Terra, prové de l’escissió de l’aigua causada pel procés fotosintetitzador.
La funció de la clorofil·la en el procés de la fotosíntesi
La clorofil·la és un pigment contingut en els cloroplasts, orgànuls específics de la cèl·lula vegetal. Per simplificar parlem de clorofil·la en general, però en realitat seria més correcte parlar de clorofil·les, en plural. A més de l’anomenada clorofil·la a, present en tots els organismes capaços de realitzar la fotosíntesi (llevat dels bacteris fotosintetitzadors), hi ha altres tipus de clorofil·les que difereixen lleugerament de la primera pel que fa a l’estructura.
Sense entrar en detalls de la descripció de la complexa molècula de la clorofil·la, donarem una ullada a les característiques químiques d’aquesta molècula, característiques que "expliquen" la seva capacitat de captar la llum i iniciar així la fotosíntesi. Convé remarcar especialment que l’anell que constitueix el "cap" de la molècula és constituït per àtoms de carboni i nitrogen units entre ells per enllaços simples i dobles alternats.
Aquests enllaços dobles amb un enllaç simple intercalat són característics de moltes altres substàncies de color i que tenen interaccions particulars amb la llum. S’anomenen enllaços dobles conjugats, i els electrons que contribueixen a formar-los, que s’uneixen als nuclis als quals pertanyen d’una manera menys estreta que els electrons de l’enllaç simple, formen un "núvol" electrònic al voltant de la molècula. Per raons físiques, la presència d’enllaços dobles conjugats confereix a la molècula capacitat per a absorbir l’energia de la llum del sol. Però aquí no acaba tot.
En efecte, l’absorció de la llum modifica l’estat energètic de la molècula. Quan la clorofil·la és colpida per un raig de llum, l’estat energètic dels electrons dels enllaços dobles resulta, per dir-ho d’alguna manera, pertorbat, i els electrons passen de l’estat energètic en què es troben normalment, anomenat estat fonamental, a un estat d’energia major, anomenat estat excitat. La diferència d’energia entre tots dos estats és, òbviament, igual a l’energia absorbida en forma de llum. Així, l’energia lumínica s’ha transformat en energia que excita els electrons. Un electró en l’estat excitat pot fer, gràcies a la quantitat addicional d’energia, el que no podria fer un electró en l’estat fonamental, és a dir, pot vèncer la força d’atracció que el vincula als nuclis dels àtoms de la molècula a la qual pertany i passar a una altra molècula. La funció de la clorofil·la durant la fase lumínica de la fotosíntesi és, doncs, iniciar tot el procés fotosintètic. La "captura" de l’energia de la llum del sol permet l’alliberament i el "transport" (a través d’una cadena de molècules) dels electrons destinats a formar molècules d’alta energia química (NADPH i ATP), que al seu torn són essencials per a l’organificació del carboni i la producció de la glucosa, processos que tenen lloc en la fase fosca de la fotosíntesi.
La fase fosca de la fotosíntesi
Abans hem vist de quina manera la fase lumínica de la fotosíntesi produïa nucleòtids reduïts (NADPH) i compostos fosforilats (ATP) d’alta energia. També ens hem pogut adonar de com i per què el procés fotosintetitzador consumeix aigua i produeix oxigen. Si tornem a fixar-nos en l’equació general de la fotosíntesi:
6CO2 + 6H2O → C6H12O6 + 6O2
ens adonem, però, que el consum d’aigua i la producció d’oxigen són tan sols una part del procés. En efecte, queda per veure com la fotosíntesi aconsegueix sintetitzar la glucosa, utilitzant el diòxid de carboni i els àtoms d’hidrogen de l’aigua (recordem que un àtom d’hidrogen té un electró i un protó). Aquesta part de la fotosíntesi és la que s’anomena fase fosca.
En aquesta fase, l’energia que ha estat acumulada en les molècules d’NADPH i ATP produïdes en la fase lumínica és utilitzada per a sintetitzar la glucosa. La fase fosca rep aquest nom perquè les seves reaccions químiques són independents de la llum. Tanmateix, també es localitza en el cloroplast en contacte estret amb la fase lumínica.
Observant l’equació general que resumeix la fotosíntesi, es podria pensar que 6 molècules de diòxid de carboni s’uneixen entre elles per a originar, en una sola reacció, un compost de 6 àtoms de carboni, com la glucosa. En química, però, una reacció simultània entre 6 molècules és un fet extremament improbable, fins i tot en els organismes vius que tenen estructures específiques capaces de captar i orientar les molècules i —gairebé es podria dir— obligar-les a actuar de la manera desitjada. En realitat, en la fase fosca, les molècules de CO2 reaccionen d’una en una. Aquesta reacció comporta l’"empaquetament" de cada molècula de CO2 en un complex orgànic preexistent, la ribulosa difosfat, derivada d’un sucre format per molècules de 5 àtoms de carboni. Amb aquesta reacció (carboxilació) el diòxid de carboni perd la naturalesa de molècula inorgànica i entra a formar part d’una molècula orgànica. D’aquesta manera és com es realitza un procés d’importància fonamental: el pas de matèria inorgànica a matèria orgànica, fenomen conegut com "organificació" del carboni.
Després de la carboxilació se succeeix una complicada cadena de reaccions (coneguda com a cicle de Calvin, pel nom del bioquímic que va contribuir al seu descobriment). Al final de la cadena tenim d’una banda 6 molècules de CO2, inicialment fixades per 6 molècules de ribulosa difosfat, reunides per a formar una molècula orgànica de 6 àtoms de carboni, precisament la glucosa, i d’altra banda les 6 molècules de ribulosa difosfat a partir de les quals havia començat el cicle i a partir de les quals pot tornar a començar.
Aquest últim resultat del cicle és tan essencial per a la vida de les plantes com la síntesi de la glucosa. Si no es formés ribulosa contínuament, la fotosíntesi s’aturaria per la manca de la molècula capaç de fixar el diòxid de carboni.
Com ja hem dit, per a portar a terme aquest cicle cal una gran despesa energètica. L’energia necessària per a sintetitzar una molècula de glucosa és una mica superior (aproximadament un terç) a la que s’obté després, quan es degrada. L’energia que cal és l’emmagatzemada durant la fase lumínica del procés fotosintètic. Això ens hauria d’ajudar a entendre del tot la importància que té per als organismes vius, i per als humans, la capacitat de les plantes (que els animals no tenen) d’absorbir l’energia del sol.
La nutrició mineral
Tret del carboni, tots els altres elements que intervenen en la constitució química dels teixits vegetals i que són necessaris per a la vida de les plantes s’obtenen del sòl. Això és vàlid tant pel que fa a l’hidrogen i l’oxigen —els dos elements que, juntament amb el carboni, constitueixen la major part del pes sec de la planta—, com a tots els altres elements que, encara que presents en quantitats menors, són igualment necessaris per a la salut de la planta.
L’absorció d’aquests elements es defineix generalment com a "nutrició mineral". Alguns dels elements absorbits (el nitrogen, el potassi, el calci, el magnesi, el fòsfor i el sofre) són necessaris en quantitats relativament elevades i per això s’anomenen macronutrients o macroelements (del terme grec "macros", gran); altres elements (el ferro, el clor, el coure, el zinc, el bor i el molibdè) són igualment indispensables per a la vida de les plantes, però en quantitats molt limitades, i per això constitueixen els anomenats micronutrients o microelements (del grec "micros", petit).
Els elements minerals estan dissolts en l’aigua del sòl en forma d’ions. En certs casos, l’àtom de l’element característic va unit a àtoms d’oxigen. És el cas del nitrogen, el fòsfor i el sofre, que s’absorbeixen en forma de nitrats, fosfats i sulfats. Són absorbits per la part apical de les arrels, que, gràcies a la seva gran superfície, aconsegueixen obtenir les substàncies minerals en quantitat suficient per a la planta, fins i tot quan la seva concentració en l’aigua del sòl és molt baixa, com passa sovint en la natura. Penseu que a vegades els ions són continguts en l’aigua del sòl en quantitats de milionèsimes de gram per litre! Un cop absorbits per les cèl·lules de l’arrel, els ions són transportats fins al cilindre central i les cèl·lules vasculars dels teixits conductors que inclouen (el xilema i el floema). Els ions continguts a l’aigua pugen pels vasos dels teixits conductors a través de les arrels i la tija fins a les fulles. Segons l’alçada i l’estructura de la planta, aquest ascens és d’uns quants centímetres o de desenes de metres.
Com pot ser que en les plantes l’aigua i les sals minerals vagin de baix a dalt, és a dir, contra la força de la gravetat? Això s’explica perquè els teixits foliars (per ser exactes, la superfície de les cèl·lules del mesofil·le foliar que són en contacte directe amb l’aire dels espais intercel·lulars) constitueixen una superfície transpirant a través de la qual s’evaporen molècules d’aigua contínuament. Aquesta pèrdua constant de molècules d’aigua al capdamunt de la columna d’aigua que té la base en les arrels, crea una mena de "força xucladora" (el terme científic és potencial hídric) que es transmet des de la superfície transpirant al llarg de tota la columna fins a l’extrem de les arrels. Per tant, les arrels tenen en tot moment la força necessària per a absorbir més aigua del sòl que substitueix l’aigua que s’ha perdut per transpiració. Les substàncies minerals dissoltes en l’aigua també són arrossegades en el flux ascendent i així poden arribar a les fulles i a totes les altres parts de la planta.
L'arrel
En una planta, la funció de l’arrel és doble: d’una banda assegura la subjecció de la planta al sòl gràcies a les seves ramificacions múltiples i, de l’altra, en proporciona l’absorció de substàncies nutritives (aigua i sals minerals).
Anatomia de l'arrel
L’arrel s’origina directament de l’embrió (arrel primària), i ben aviat es ramifica i produeix arrels secundàries que al seu torn es poden ramificar per a produir arrels terciàries, i així successivament. Tot plegat constitueix el sistema radical axonomorf, el desenvolupament i la complexitat del qual varien segons l’espècie i segons les condicions edàfiques i climàtiques a les quals estigui sotmesa la planta. Perquè la planta gaudeixi de bona salut cal que les arrels puguin assolir un desenvolupament i una ramificació suficients, de manera que puguin oposar una resistència adequada al desarrelament (originat pel vent o els animals, etc.) i augmentar al màxim la superfície útil d’absorció dels nutrients del sòl. Les modalitats de ramificació i desenvolupament del sistema radical no són iguals en totes les plantes. Esquemàticament, les arrels poden adoptar dues configuracions principals: la napiforme i la fasciculada. En la configuració napiforme l’arrel primària és molt més desenvolupada que les secundàries; en la configuració fasciculada, en canvi, l’arrel primària deixa de créixer d’una manera prematura mentre que les arrels secundàries, les terciàries, etc. assoleixen un gran desenvolupament. El que no varia és l’estructura anatòmica interna de l’arrel. Cada ramificació repeteix l’estructura de l’arrel original, de manera que n’hi ha prou d’estudiar aquesta estructura per a conèixer l’anatomia de tot el sistema radical.
L’extrem de cada ramificació de l’arrel s’anomena àpex radicular. És recobert per una mena de funda protectora, anomenada còfia o pilorriza, que serveix per a evitar el contacte directe de l’àpex jove i delicat amb les aspres partícules del sòl que el perjudicarien. Aquest contacte es fa a través de la còfia, constituïda per un nombre reduït de capes cel·lulars. Les capes més exteriors, a causa del fregament amb el sòl es desgasten contínuament i són substituïdes per les cèl·lules de sota.
Perquè la còfia no es consumeixi cal, òbviament, que hi hagi un subministrament continuat de cèl·lules noves. D’aquest subministrament se n’ocupa la zona pilífera de l’àpex, que es diferencia de la còfia i dels següents trams d’arrel per les característiques peculiars de les seves cèl·lules. Són cèl·lules meristemàtiques o embrionàries típiques, la principal activitat de les quals és la divisió. Les divisions segueixen un pla sobretot perpendicular a l’eix de l’arrel, de manera que una de les dues cèl·lules resultants de cada divisió estarà sempre més desplaçada cap a l’àpex i l’altra més cap a la base. Les cèl·lules que progressivament van quedant lluny de l’àpex, empeses per les noves que provenen de les divisions cel·lulars que se segueixen produint, perden la capacitat de dividir-se i es van diversificant. Assumeixen progressivament, a partir d’un procés de diferenciació, l’aspecte dels diversos tipus cel·lulars que constitueixen la part adulta de la planta.
Molt a prop de l’àpex de l’arrel hi ha les anomenades zones de creixement i diferenciació, on les cèl·lules creixen per elongació i al mateix temps es diferencien adquirint les característiques típiques del teixit del qual formaran part.
Modificacions de les arrels
En moltes plantes les arrels poden adoptar altres funcions a més de les normals. Sovint, les arrels verticals s’engruixeixen i esdevenen òrgans de reserva de la planta; la pastanaga, el nap i la remolatxa en són casos típics. Aquestes arrels s’anomenen napiformes, i són àmpliament emprades en l’alimentació humana pel seu contingut de substàncies energètiques.
En altres casos, les arrels poden adoptar una funció reguladora de l’alçària de la planta respecte al sòl. És el que passa per exemple en les plantes amb disposició en roseta o en les plantes amb bulbs o rizomes (vegeu "Modificacions de la tija"), que durant l’estació desfavorable perden la part aèria. Al final del període vegetatiu les arrels es contreuen i provoquen la retirada de la gemma apical una mica per sota de la superfície del sòl de manera que quedi protegida.
En algunes plantes que viuen en ambients aquàtics, les arrels també poden encarregar-se de la tasca de sosteniment del tronc per sobre de la superfície de l’aigua o de desenvolupar arrels respiratòries que sobresurten de l’aigua per airejar les parts submergides (els manglars).
Un altre tipus d’arrels especials són les anomenades caulogèniques, que es desenvolupen directament a partir del tronc o de les fulles. En moltes gramínies, com el blat de moro, els nusos inferiors de la tija produeixen en tot el seu diàmetre nombroses arrels caulogèniques que substitueixen l’arrel principal, la qual interromp d’una manera prematura el seu desenvolupament. Les plantes amb tiges reptants (els estolons) desenvolupen a l’altura dels nusos i cap avall arrels adventícies i cap amunt brots; és el cas típic de la maduixera. Moltes plantes enfiladisses, com l’heura, produeixen arrels caulogèniques (en aquest cas s’anomenen adherents o garfis) que els permeten adherir-se a les tiges d’altres plantes o a substrats de diversa mena. És interessant el cas de les arrels paliformes o fúlcries del Ficus religiosa, que neixen de les branques i es dirigeixen cap a terra, mentre la seva part aèria fa la funció de sosteniment. A conseqüència de lesions o ruptures de les branques també creixen arrels adventícies. Aquest fenomen és àmpliament explotat per l’agricultura per a obtenir noves plantes a partir de brots (propagació per estaca).
L'estructura primària
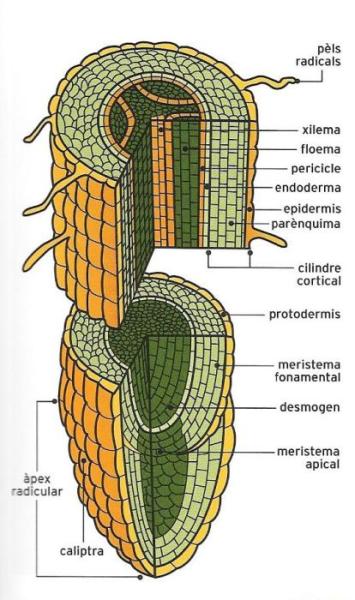
Esquema de l'estructura de l'arrel d'una planta. La meitat superior representa la zona pilífera, on els pèls radicals, prolongacions molt fines i tendres de les cèl·lules de la rizodermis, s'encarreguen de l'absorció. A la meitat inferior i terminal és on el meristema produeix les noves cèl·lules que permetran el creixement en longitud. A l'àpex, les cèl·lules suberificades protegeixen l'arrel contra el fregament amb el sòl.
ECSA
A una distància d’uns quants centímetres de l’àpex els teixits ja s’han diferenciat completament i es comencen a organitzar per a originar "l’estructura primària". En l’estructura primària de l’arrel es distingeixen tres regions que, de fora endins, són: l’epidermis, el còrtex i el cilindre central.
L’epidermis, com veurem més endavant ("Els teixits tegumentaris"), és un típic teixit tegumentari o de revestiment, constituït per una sola capa de cèl·lules, disposades regularment les unes al costat de les altres de manera que no deixen espais intercel·lulars. L’epidermis és proveïda d’uns filaments exteriors característics, els pèls radicals. Amb el desenvolupament dels pèls radicals, la planta aconsegueix l’important objectiu d’augmentar la superfície de contacte amb l’exterior i, per tant, la seva capacitat d’absorció de l’aigua continguda al sòl i de les sals minerals dissoltes en l’aigua. Efectivament, la funció principal d’aquesta zona de l’arrel és l’absorció, tot i que l’epidermis també té una funció protectora.
Els pèls radicals, i tota l’epidermis, tenen una vida curta: al cap de pocs dies, els pèls i les cèl·lules més velles moren i es desprenen de l’arrel. Mentrestant, i gairebé al mateix ritme, a l’àpex es formen altres pèls a mesura que l’arrel es va allargant. Per tant, la llargada i la posició de la zona pilífera es mantenen més o menys constants.
A les zones on l’epidermis desapareix, la funció protectora és assumida per la capa cel·lular de sota, que es modifica per tal de poder realitzar-la. Això s’aconsegueix mitjançant la suberització o la cutinització de la paret cel·lular, a la qual ja hem fet referència a "La paret cel·lular". Les cèl·lules modificades d’aquesta manera constitueixen l’exoderma, que és impermeable a les substàncies dissoltes al sòl, de manera que a partir d’aquesta zona l’arrel perd la capacitat d’absorció i, per tant, tan sols conserva la funció de fixació de l’òrgan al substrat.
Sota l’epidermis i/o l’exoderma hi ha el còrtex, que és constituït per un nombre de capes cel·lulars variable segons les espècies. Es tracta preferentment de cèl·lules de tipus parenquimàtic, de forma arrodonida o polièdrica, sovint allargades en sentit longitudinal, i que només es toquen per la superfície. Per tant, es forma un sistema d’espais intercel·lulars. La capa més interna de l’escorça, l’endoderma, constitueix una mena de filtre, que té la capacitat de distingir quines de les substàncies absorbides són útils per a la planta (i per tant les deixa penetrar en el cilindre central de l’arrel perquè siguin distribuïdes a tota la planta), i quines són perjudicials o simplement inútils (les quals no deixa passar).
La capa més externa del cilindre central és el pericicle, normalment constituït per una sola capa de cèl·lules, a partir del qual s’originen les arrels laterals. A l’interior del pericicle hi ha els teixits conductors de l’arrel, és a dir, els sistemes "conductors" que permeten transportar la saba a totes les parts de la planta.
L'estructura secundària
Arriba un moment en què la capacitat de creixement dels teixits que formen el cos primari, capacitat que rau essencialment en la distensió de les cèl·lules i en el consegüent augment de volum, impedeix que l’arrel desenvolupi amb èxit les seves funcions. En una planta on la part aèria (la tija i les fulles) també s’està desenvolupant i per tant necessita un subministrament apropiat d’aigua i sals minerals, és encara més clar. Aleshores, algunes cèl·lules del cos primari de l’arrel tornen a adquirir característiques i capacitats meristemàtiques. Es comencen a dividir i així originen el cos secundari de l’arrel.
El creixement secundari és un procés extremament complicat que aquí no descriurem amb detall; de tota manera, en tornarem a parlar a propòsit de la tija. Aquí ens limitarem a posar en relleu que els teixits responsables d’aquest creixement, els càmbiums (o meristemes laterals), formen dos anells continus (anells que en realitat són beines cilíndriques). A través de la divisió cel·lular poden, per tant, augmentar de gruix indefinidament, mentre la planta visqui. També són capaços d’anar augmentant progressivament la seva circumferència. No totes les plantes presenten creixement secundari, cosa que tant és vàlida per a l’arrel com per a la tija. No el presenten la majoria de les monocotiledònies, i en algunes dicotiledònies herbàcies de cicle anual a penes s’insinua. En canvi, en la majoria de les dicotiledònies i en totes les gimnospermes el creixement secundari és ben present. Un cas particular és el d’algunes monocotiledònies, en què el creixement secundari de l’arrel només afecta el còrtex, mentre que el cilindre central roman en l’estadi primari.
La tija
La part de la planta que normalment es desenvolupa per sobre del sòl constitueix el brot, format per la tija i les fulles, ben diferents quant a morfologia i funció, però estretament interconnectades perquè provenen del mateix àpex vegetatiu. La tija forma l’eix del brot i s’uneix a l’arrel directament des de la base. A més d’aguantar les fulles i, per tant, fer una funció mecànica, actua de punt d’unió entre les diverses parts de la planta i ho fa a través dels complexos sistemes de teixits conductors, alguns destinats al transport de l’aigua i les sals minerals absorbides per l’arrel i uns altres encarregats del transport de les substàncies orgàniques produïdes per les fulles.
Anatomia de la tija
L’àpex de la tija s’anomena gemma. Com l’àpex radicular, inclou els meristemes primaris o apicals, dels quals provenen, directament o indirectament, tots els altres teixits. Ara bé, la tija presenta una diferència important respecte de l’arrel; mentre que a partir de l’arrel principal només es poden desenvolupar altres arrels, a la tija també es desenvolupen apèndixs laterals amb estructures diverses que tenen una funció importantíssima per a l’organisme vegetal: les fulles. Neixen en zones particulars de la tija anomenades nusos. En alguns casos, d’aquests nusos surten gemmes amb les mateixes característiques que l’àpex i que en créixer originen cadascuna una ramificació. Així es determina el "port" de la planta, que és característic de cada espècie. Els trams de tija compresos entre dos nusos s’anomenen entrenusos.
La tija pot adoptar conformacions i dimensions molt diverses. En les plantes de cicle vital curt, anual o biennal, és normalment herbàcia, prima i de dimensions modestes, mentre que en les plantes perennes, que poden arribar a viure centenars o fins i tot milers d’anys, és lignificada i revestida per una escorça suberosa (aleshores es parla de tronc). La conformació és arbustiva quan la ramificació del tronc té lloc a la base, i és arbòria quan la ramificació comença més amunt. Els arbusts tenen dimensions més aviat limitades i com a màxim assoleixen uns quants metres d’alçada. Els arbres, en canvi, són molt més alts i en alguns casos (les sequoies i els eucaliptus) presenten dimensions veritablement excepcionals i poden superar els cent metres. Òbviament, els troncs d’aquestes dimensions tenen una base de molts metres de diàmetre.
Es distingeixen dos tipus generals de ramificació de la tija: la monopòdica i la simpòdica. En la primera, l’eix principal continua creixent indefinidament per acció de l’àpex vegetatiu primari, i les branques laterals mai no superen les dimensions de l’eix principal. És el cas de moltes coníferes com l’avet o de dicotiledònies com l’auró. En la ramificació simpòdica l’àpex vegetatiu primari deixa de créixer prematurament i és substituït per una gemma lateral que, al seu torn, té un període de desenvolupament limitat i també és substituïda, i així successivament. El resultat és una capçada espessa i molt ramificada. Un exemple típic de ramificació simpòdica és el de l’alzina o el bedoll, encara que en les parts adultes de la planta que han augmentat de gruix no és fàcil de reconèixer.
Després de descriure l’estructura exterior i el port de la tija, n’estudiarem l’estructura interior. A partir de l’àpex, es dóna la mateixa successió que hem vist en l’arrel. A sota de la zona meristemàtica trobem les zones de determinació, de diferenciació, d’estructura primària i d’estructura secundària. Pot ser que aquesta darrera no aparegui ni a la tija ni a les arrels, ja que no totes les plantes presenten creixement secundari. La llargada de cadascuna d’aquestes zones pot variar molt: d’unes quantes centèsimes de mil·límetre en els meristemes apicals i en la zona de determinació a uns quants centímetres en la zona de diferenciació; dels molts centímetres de l’estructura primària a les moltes desenes de metres a les quals es desenvolupa el cos secundari de les plantes anomenades "de tronc alt".
Les característiques, la disposició i l’activitat de les cèl·lules de les tres primeres zones (l’apical, la de determinació i la de diferenciació) són iguals que les descrites per a les arrels. L’àpex de la tija es distingeix del de les arrels per l’absència de la còfia, perquè, com que es desenvolupa fora del sòl, no li cal una estructura de protecció.
Modificacions de la tija
Respecte de la seva conformació típica, la tija pot presentar modificacions que s’anomenen metamorfosis. Són característiques de cada espècie i estan relacionades amb especialitzacions funcionals i ecològiques. Algunes plantes poden desenvolupar tiges reptants subterrànies, els rizomes, de creixement horitzontal, amb fulles reduïdes a esquames d’on surten brots aeris erectes. De vegades el rizoma és molt ramificat, com en el cas del gram, en què un sol individu es pot estendre per superfícies de sòl relativament extenses, o bé pot ser curt i engruixit amb abundants parènquimes de reserva. Quan les tiges reptants es desenvolupen a la superfície del sòl es parla d’estolons (vegeu "Modificacions de les arrels").
Les plantes que viuen en climes secs (xeròfiles) poden formar tiges aplanades verdes (els cladodis) que adopten la funció i, de vegades, fins i tot la forma de les fulles (el galzeran), mentre les fulles veritables són reduïdes a esquames o transformades en espines o cauen abans d’hora. En certes plantes suculentes, com per exemple la figuera de moro, es formen cladodis de grans dimensions, mentre que en altres la tija esdevé globulosa (certes cactàcies, euforbiàcies i altres) o cilíndrica i de gran diàmetre.
Les tiges aèries, com les fulles, poden transformar-se en espines, simples o de vegades també ramificades. Aquesta modificació és molt freqüent en les plantes xeròfiles, però també en altres, i la seva funció és protegir la planta dels animals. Les espines de l’arç blanc són constituïdes per curts brots axil·lars simples i curts, mentre que les espines de l’arç de tanques són llargues i ramificades.
En les plantes enfiladisses les tiges aèries, però sovint també les fulles, es poden transformar en caulicles simples o ramificats, formats per apèndixs prims i allargats, capaços d’enrotllar-se sobre branques o altres suports. A vegades, l’extrem dels caulicles pot formar discs adhesius que es fixen al substrat; és el cas del cep del Canadà (aleshores es parla de circells caulinars).
Una altra modificació freqüent de la tija, sovint present en les tiges subterrànies, són els tubercles caulinars, que s’han format per l’engruiximent de parts determinades (els tubercles), i que tenen com a funció l’acumulació de material de reserva. Un cas típic és la patata. Algunes parts de la tija convertides en tubercles poden transformar-se en òrgans perennes, en plantes que perden la part aèria durant l’estació desfavorable, com per exemple en el còlquic. Quan tota una gemma esdevé òrgan subterrani sovint es forma un bulb, amb l’eix molt curt i engruixit i amb les fulles transformades en esquames carnoses que assumeixen la funció de reserva i protegeixen l’àpex vegetatiu.
L'estructura primària
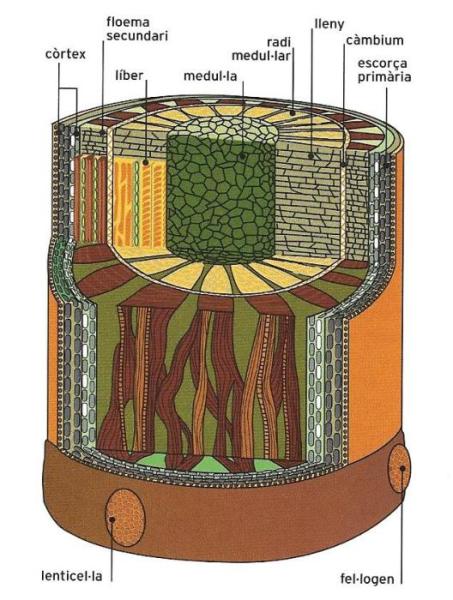
Esquema de l'estructura primària i secundària de la tija. L'estructura primària és igual a la de l'arrel. La secundària, característica sobretot de les plantes llenyoses, es deu a l'activitat del càmbium, que forma un anell de cèl·lules capaces de produir noves cèl·lules del xilema cap endins, el lleny, i noves cèl·lules del floema cap enfora, el líber
ECSA
Si observem al microscopi una secció transversal de la tija jove d’una planta a uns centímetres per sota de l’àpex, veurem una estructura més aviat complexa en la qual ja hi ha diferenciats nombrosos teixits. La secció que examinem no és sinó l’estructura primària de la tija, on de la mateixa manera que a l’arrel es poden distingir tres capes concèntriques principals: l’epidermis, el còrtex i el cilindre central (o estela). Si comparem la secció transversal d’una tija d’estructura primària i la d’una arrel, de seguida s’evidencia una diferència: ambdós òrgans presenten un desenvolupament diferent del còrtex i el cilindre central. En la tija el gruix del còrtex és reduït i el del cilindre central és més desenvolupat, mentre que a l’arrel les capes corticals ocupen la major part del diàmetre i el cilindre central és relativament petit.
En examinar cada capa i cada teixit, veiem que l’epidermis de la tija és constituïda per una capa (rarament més d’una) de cèl·lules més aviat aplanades i allargades en el sentit longitudinal, disposades molt juntes de manera que no quedin espais intercel·lulars. La paret cel·lular és una prima paret primària constituïda per cel·lulosa, excepte en la cara que dóna a l’exterior, en contacte directe amb l’atmosfera. En aquesta cara la paret sempre apareix cutinitzada i, a més, pot presentar cutícules i revestiments cerosos externs. Evidentment, totes aquestes característiques tenen la funció de conferir a l’epidermis, i per tant a la tija, una elevada capacitat protectora, impermeabilitzant, aïllant i antitranspirant.
Per sota de l’epidermis hi ha l’escorça de la tija. Per bé que proporcionalment és menys extensa que la de l’arrel, realitza força funcions. S’hi troben dos tipus de cèl·lules: les parenquimàtiques i les de sosteniment. En les tiges joves, les cèl·lules parenquimàtiques de les capes corticals més externes contenen gairebé sempre cloroplasts, tal com podem constatar amb facilitat observant-ne el color verd. Aquestes cèl·lules realitzen, doncs, la fotosíntesi clorofíl·lica —tot i que aquesta funció és típica de les fulles i la que es porta a terme a la tija només és una part petita de la fotosíntesi total de la planta—. Anant cap a les capes més internes, i per tant menys exposades a la llum, veiem que els cloroplasts són substituïts per amiloplasts, que acumulen el midó obtingut a partir de la fotosíntesi de les capes més externes, o el que prové de les fulles a través del teixit conductor. Així doncs, els amiloplasts actuen de teixit de reserva.
La funció de sosteniment de l’escorça és competència del teixit mecànic o de sosteniment, format pel colènquima i l’esclerènquima. Les parets cel·lulars del colènquima són parcialment engruixides, mentre que les de l’esclerènquima són engruixides del tot. La capa més interna del còrtex de la tija es caracteritza sovint per l’alt contingut de midó, per la qual cosa rep el nom de beina amilàcia.
El cilindre central de la tija presenta una estructura força complexa que, a més, no sempre és la mateixa en totes les plantes. En efecte, mentre que l’arrel presenta una uniformitat bàsica en la disposició dels feixos vasculars, i en la tija la posició del xilema i el floema és invariable —tenen una disposició col·lateral, amb el floema cap enfora i el xilema cap endins—, la disposició dels feixos dins el cilindre no és sempre la mateixa. L’estructura més comuna, present en totes les gimnospermes i en moltes dicotiledònies, és l’anomenada euestela: els feixos conductors es distribueixen en una corona regular desplaçada cap a la part exterior del cilindre central i formen així un anell discontinu que s’inclou en el teixit parenquimàtic. Aquest darrer teixit es divideix en: pericicle (les capes externes dels feixos conductors), que en alguns casos és molt reduït o fins i tot absent, medul·la (la part interna dels feixos conductors) i radis medul·lars (la part que va d’un feix a un altre).
L'estructura secundària
L’estructura euestèlica del cilindre central, a més de ser la més extensa entre les plantes superiors, és també aquella que més típicament origina l’estructura secundària de la tija. Les plantes que tenen altres tipus d’estructura del cilindre central no presenten creixement secundari, i si el presenten és molt limitat. En aquestes plantes (les monocotiledònies i part de les dicotiledònies) l’estructura primària és, doncs, la definitiva. Contràriament, en les plantes euestèliques l’estructura primària només apareix a les parts joves, que a mesura que es desenvolupen i envelleixen adopten progressivament l’estructura secundària.
En parlar de l’arrel ja hem examinat les raons que motiven el creixement secundari. És possible gràcies a l’activitat d’unes cèl·lules especials: les cèl·lules meristemàtiques secundàries o laterals. El teixit que formen aquestes cèl·lules, anomenat càmbium, s’origina o bé a partir de les cèl·lules centrals del xilema i el floema o bé de les cèl·lules dels radis medul·lars. Aquestes dues parts del càmbium, en unir-se, formen una mena d’anell continu que recorre tota la tija. Quan les cèl·lules del càmbium es divideixen, com en totes les cèl·lules meristemàtiques, s’originen noves cèl·lules del xilema cap endins i noves cèl·lules del floema cap enfora, responsables del creixement en gruix de la tija. A més del càmbium, hi ha un altre teixit meristemàtic secundari que participa, per bé que en menor mesura, del creixement secundari de la tija: el fel·logen. Neix del còrtex de la tija i també forma un anell continu. Quan es divideix produeix suro nou i el teixit de protecció que revesteix la tija.
Els anells de creixement
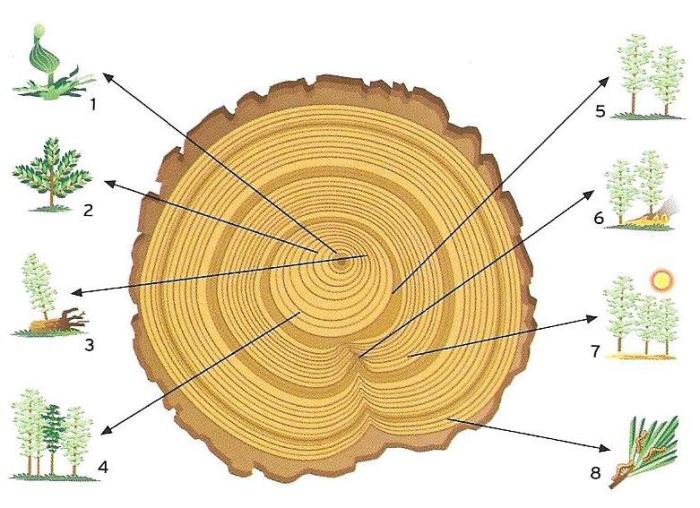
Esquema de la seqüència d'esdeveniments enregistrats en els anells d'un pi des del seu naixement fins que mor al cap d'uns 60 anys.
Primer anell format l'any de la germinació (1). El cop que rep d'un arbre veí que ha caigut (3) i l'efecte d'un incendi (6) deixen una cicatriu en els anells corresponents. La competència amb altres arbres veïns per la llum, l'aigua i els nutrients (4), els períodes de sequera (7) i els atacs de processionària (8) queden gravats en els anells més estrets. Per contra, quan els arbres amb desavantatges moren, i per tat, es produeix un alliberament de recursos que els altres aprofiten (5) i quan no hi ha competència per l'espai (2), el seu creixement és més ràpid i els anells són amples.
ECSA
La majoria d’arbres i espècies llenyoses de les zones temperades i boreals aturen el seu creixement durant l’època desfavorable de tardor i hivern i el reprenen quan les condicions climàtiques tornen a ser favorables, a la primavera i a l’estiu. Aquest patró anual d’activitat i repòs queda reflectit en l’estructura de fusta del tronc, de les branques i de les arrels, formada per capes concèntriques que en un tall transversal es veuen com uns anells. En moltes espècies mediterrànies, com l’alzina, el garric, el llentiscle o el pi blanc, malgrat que es veuen anells de creixement, aquests no acostumen a ser anuals a causa de l’elevada variabilitat climàtica intraanual. Aquestes espècies poden créixer tot l’any amb aturades més o menys llargues a qualsevol època quan hi ha manca d’aigua o les temperatures són baixes, la qual cosa denota l’origen tropical-subtropical de moltes d’elles.
Els anells de creixement es formen a partir d’un teixit meristemàtic secundari, el càmbium, situat entre l’escorça i la fusta. El càmbium genera xilema o fusta cap a l’interior i floema cap a l’exterior i té l’habilitat de no quedar-se atrapat sota el xilema general, és per això que l’últim anell format és el més extern. La formació d’un anell és ràpida al començament i s’alenteix a mesura que avança l’estiu. Aquestes diferències de velocitat també queden reflectides en les característiques de la fusta que els constitueix i per aquesta raó en cada anell es distingeixen dues bandes. La fusta primerenca, formada al principi del període de creixement, té les cèl·lules més grans i la paret cel·lular més prima. Però a mesura que l’anell es va formant, les cèl·lules es fan cada cop més petites i la seva paret cel·lular més gruixuda, cosa que dóna lloc a la fusta tardana. En moltes espècies, principalment en les coníferes, la fusta tardana és també d’un color més fosc.
La transició entre la fusta tardana d’un anell i la primerenca de l’anell de l’any següent és brusca; aquesta juxtaposició permet distingir els anells formats cada any. Comptant els anells s’ha comprovat que alguns arbres poden viure fins a 4 000 anys, com és el cas del Pinus longaeva als Estats Units o del Fitzroya cupressoides, de més de 3 600 anys, a Xile. A Espanya, els arbres vius més vells pertanyen a Pinus pinea, d’uns 1 000 anys (Andalusia), i Pinus uncinata, d’uns 600 anys (Catalunya).
L’activitat del càmbium és molt sensible a les condicions de l’entorn i en els anells queden enregistrats tots aquells esdeveniments que han afectat l’arbre al llarg de la seva vida. Per altra banda, els arbres són els organismes que viuen més anys, i aquesta informació, que pot ésser recuperada i convenientment analitzada, permet estudiar processos ecològics i ambientals amb una resolució anual. Els anells de creixement són sensibles a les variacions del clima, la contaminació atmosfèrica, les característiques físiques i químiques del sòl, la competència amb els veïns, els efectes dels animals que s’alimenten de les fulles, les pertorbacions, etc. Tots aquests factors modifiquen el nombre i la mida de les cèl·lules, cosa que es reflecteix en les taxes d’engruiximent i en les característiques físiques i químiques dels anells.
Altres factors, com, per exemple, les ferides produïdes pel foc, l’home, els animals, els cops de pedra i dels arbres veïns que cauen, afecten el càmbium i queden gravats en forma de cicatriu que marca l’anell de l’any corresponent.
La ciència que es dedica a obtenir i analitzar aquesta informació és la dendrocronologia, que es basa en la datació correcta dels anells —assignant a casdascun d’ells l’any en què es va formar— i en l’anàlisi dels patrons temporals de les sèries d’anells establint-ne cronologies. La dendrocronologia té moltes aplicacions en diferents camps de recerca.
L’anàlisi dels anells permet analitzar la dinàmica de les poblacions i reconstruir la història dels boscos, les variables que els caracteritzen, el règim de pertorbacions com gelades, vents, tales, incendis o les activitats dels animals que els han afectat al llarg del temps.
La dendrocronologia s’aplica també a l’estudi dels canvis de cabal dels rius i del nivell de les capes freàtiques i, en geomorfologia, per analitzar els moviments del sòl.
Històricament, s’ha aplicat a l’estudi del clima actual i del passat (dendroclimatologia). Com que la variació anual del clima afecta tots els arbres d’una regió d’una manera similar, les seqüències d’anells de diferents arbres s’assemblen, presenten una sincronia. Per exemple, els anys climàticament bons queden enregistrats amb anells més gruixuts, mentre que els anys amb un clima més extrem queden enregistrats amb anells més estrets o amb d’altres característiques diferenciades. Així, podem reconèixer en qualsevol arbre l’anell dels anys 1984 i 1986, que són més estrets a causa de la sequera, els del 1931 i del 1963 pel fred i els del 1871 per glaçades tardanes a la primavera. Això serveix de base per a reconstruir el clima del passat i veure les variacions que ha sofert. Al centre d’Europa i a Irlanda s’han establert cronologies de més de 7 000 anys. Aquest fet ha permès estudiar les fluctuacions del clima en aquest període i, al mateix temps, datar amb precisió cases, catedrals, obres d’art i fins i tot construccions prehistòriques.
La fulla
La fulla és el tercer i darrer òrgan en què s’estructura el "cos" de les plantes superiors. Li pertoquen funcions fonamentals tant per a la vida vegetativa de la planta com per a la reproducció. Els aparells reproductors de les plantes superiors, les flors, no són sinó fulles, per bé que modificades fins al punt que no presenten la més mínima semblança amb les fulles pròpiament dites.
Anatomia de la fulla
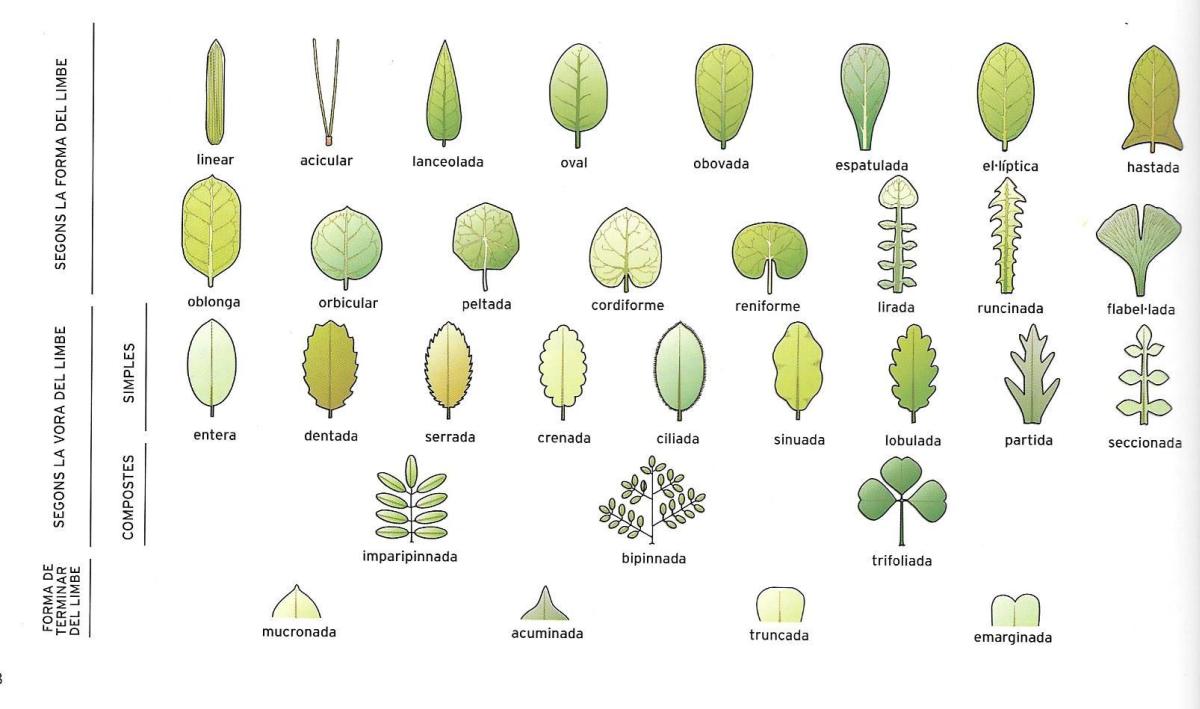
La gran diversitat de formes que poden presentar les fulles es deu a les variacions que prenen cadascuna de les seves parts. El limbe, verd, laminar i sovint molt prim, varia molt de forma, però també de tipus de nervadures i de graus de divisió. La base del limbe pot presentar així mateix formes diferents, i en alguns casos fa com una beina que embolcalla la tija, o té una mena de petites fulletes acompanyants, les estípules. El pecíol, igualment, pot presentar-se modificat en forma d'ales, o extremadament reduït.
ECSA
La fulla és, almenys en la majoria dels casos, un òrgan asimètric. En efecte, a primer cop d’ull es pot observar una diferència (quant al color, el relleu de les nervadures, etc.) entre la cara superior, és a dir, l’orientada cap amunt i per tant exposada directament a la llum solar, i la cara inferior, orientada cap avall. En la majoria de les plantes la fulla és, per tant, un òrgan bifacial. Les fulles bifacials formen un angle gairebé recte amb la tija en la qual s’insereixen, a diferència de les equifacials, que formen un angle agut.
Una fulla típica completament desenvolupada és constituïda per tres parts: 1) una beina foliar, que cobreix parcialment o totalment la tija de la qual surt la fulla, 2) una part allargada o pecíol i 3) una part ampla i plana anomenada làmina foliar o limbe. Aquest és un esquema bàsic en el qual hi pot haver infinitat de variacions.
En efecte, la fulla descrita és la pròpia de les planifòlies (és a dir, les plantes de fulles amples), però la majoria de les fulles de les gimnospermes tenen forma d’agulla (per això s’anomenen aciculifòlies) i per tant no presenten la divisió en beina, pecíol i limbe. Les planifòlies, amb tot, també poden estar mancades d’alguna d’aquestes tres parts: sovint és el pecíol que hi falta (aquestes fulles que s’insereixen a la tija directament pel limbe s’anomenen "fulles sèssils"); pot ser que el desenvolupament de la seva beina sigui molt diferent i assoleixi dimensions semblants a les del limbe (com en les gramínies) o pot ser que la beina es redueixi a un petit apèndix a la base del limbe o el pecíol.
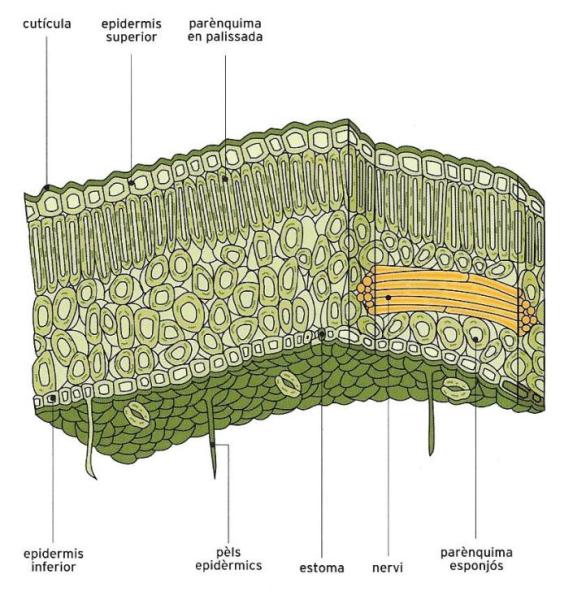
Tall transversal del limbe d'una fulla bifacial, amb la cara superior ben cutinitzada, preparada per a captar la radiació solar, i la inferior més adequada per a fer l'intercanvi de gasos i aigua amb l'exterior. El teixit intern, el mesofil·le, es diferencia en dues parts: la més propera a l'anvers realitza preferentment la fotosíntesi i és més gruixuda com és sol reben les fulles; la més pròxima al revers, al seu torn, s'ocupa de captar i distribuir el diòxid de carboni.
ECSA
Una propietat de la fulla que la distingeix de la tija i de l’arrel és el fet de presentar un creixement definit, al contrari d’aquests dos òrgans, que van creixent indefinidament mentre la planta viu. Efectivament, al cap d’un temps la fulla deixa de créixer, i com a conseqüència els teixits que la constitueixen degeneren i moren progressivament, i la fulla es desprèn de la planta.
Aquest fet passa tant en les plantes on el creixement i la caiguda de les fulles són fenòmens estacionals (les plantes de fulla caduca o caducifòlies), com les alzines, els faigs, els plàtans, els aurons, etc., com en aquelles on aquests fenòmens no són estacionals sinó continus (plantes perennifòlies), com els pins, els avets, els garrics, l’heura, etc. Hi ha, però, una diferència: mentre que les fulles de les primeres surten i es desenvolupen en un període determinat i gairebé limitat de l’any (la primavera) i s’assequen i cauen en un altre període igualment limitat (la tardor), les fulles de les segones experimenten aquests fenòmens al llarg de tot l’any i d’una manera no gaire visible. Certament, no s’ha de creure que les perennifòlies conserven les mateixes fulles durant tota la vida. El que passa és que el ritme de formació de les fulles noves és el mateix que el de caiguda de les fulles velles, de manera que l’aspecte global de la planta no canvia. D’altra banda, si s’observa el gran nombre d’agulles mortes o de fulles seques normalment presents als peus dels pins, dels avets o de les planifòlies perennifòlies com les magnòlies i els llorers, es fàcil constatar que a les perennifòlies també se’ls assequen i els cauen les fulles.
La funció fonamental de la fulla és portar a terme la fotosíntesi clorofíl·lica. Amb aquesta finalitat, la fulla requereix tres factors fonamentals: el diòxid de carboni (CO2), present en l’atmosfera, l’aigua, absorbida majoritàriament del sòl per les arrels, i les radiacions solars, que proporcionen l’energia. En la fulla, tot està destinat a garantir de la manera més eficaç possible l’obtenció d’aquests tres elements indispensables per a la vida de la planta. Per a aconseguir-ho la relació entre la superfície i el volum de la fulla ha de ser elevada.
La fulla, que tendeix a ser plana i sovint té una superfície d’uns quants centímetres quadrats, té normalment un gruix de pocs mil·límetres. Això li permet captar gran quantitat de la radiació solar i també de diòxid de carboni. L’elevada relació superfície/volum, evident en les planifòlies, pot ser menys aparent en les fulles aciculiformes típiques de les coníferes, però en aquestes plantes també és considerable. A més, les fulles aciculiformes responen a algunes exigències de subministrament hídric i de defensa contra el glaç que les coníferes han de satisfer per a suportar el clima, en general més aviat sever, de les zones on viuen.
És, doncs, indispensable una superfície àmplia per a absorbir el diòxid de carboni i la llum necessaris per a la fotosíntesi. Això no obstant, aquesta superfície comporta un risc molt perillós per a la fulla exposada i la planta en conjunt: la pèrdua d’aigua per transpiració. L’aigua absorbida del sòl circula per les arrels i la tija fins a les fulles. Aquí topa amb una capa prima, exposada de ple a la calor solar i al vent i, per tant, està en les millors condicions per a passar de l’estat líquid al de vapor, travessar la superfície foliar i perdre’s en l’atmosfera. Si l’aigua li manqués, la fulla es quedaria sense el component indispensable per a viure i alhora sense el protagonista principal de les reaccions químiques de la fotosíntesi. Més endavant veurem de quina manera l’estructura interna de la fulla la protegeix d’aquest perill. Així, doncs, massa transpiració mata la planta perquè s’acaba l’aigua, però poca transpiració també la mata perquè s’atura la circulació. S’ha d’arribar a un punt d’equilibri.
En examinar, en una secció transversal, l’estructura d’una fulla bifacial s’observen, a partir de la cara superior, quatre capes successives: l’epidermis superior, el teixit en palissada, el teixit lacunar i el mesofil·le foliar.
L’epidermis superior està revestida de cutícula. Generalment és constituïda per un sol feix de cèl·lules en les plantes que viuen en climes temperats o humits, i per diversos estrats en les plantes que viuen en climes calorosos i àrids, ja que així els protegeix millor de la pèrdua excessiva d’aigua per transpiració. Per sota del nivell epidèrmic hi ha el teixit en palissada. És format per cèl·lules allargades, disposades amb molta regularitat les unes al costat de les altres, amb pocs espais intercel·lulars, la qual cosa confereix al teixit l’aspecte de palissada. Aquest teixit pot ser uniestratificat o pluriestratificat, no solament segons l’espècie sinó també, en una mateixa planta, segons la posició de la fulla i la quantitat de llum que rep. En plantes de tronc alt, com per exemple el faig, s’observa que les fulles situades a la part exterior de la capçada, i per tant exposades directament a la llum solar, són en conjunt més gruixudes i tenen un teixit en palissada ben desenvolupat i pluriestratificat. En canvi, les fulles situades a l’interior de la capçada, que queden a l’ombra, són molt més primes i el teixit en palissada és uniestratificat i menys desenvolupat.
Per sota del teixit de palissada apareix un teixit constituït per cèl·lules de forma molt diversa i entre les quals queden amplis espais intercel·lulars. Es tracta del teixit lacunar, que, juntament amb el teixit en palissada, constitueix el mesofil·le foliar, o sigui el teixit encarregat de la fotosíntesi clorofíl·lica (el parènquima clorofíl·lic). Les cèl·lules del mesofil·le sempre contenen abundants cloroplasts, els orgànuls cel·lulars on es produeix la fotosíntesi clorofíl·lica. En realitat, pel que fa a la fotosíntesi, el teixit en palissada és molt més important que el lacunar (efectivament, aquestes cèl·lules contenen entre el 70 i el 85% dels cloroplasts totals de la fulla)i és el responsable principal de l’absorció i la transformació de l’energia lumínica.
El teixit lacunar, però, gràcies a la seva estructura particular, també té una funció fonamental, ja que a través seu es produeix l’absorció del diòxid de carboni i la seva distribució a totes les cèl·lules del mesofil·le.
Els feixos vasculars, que des de la tija es ramifiquen vers cada fulla, a la qual aporten les sals minerals i l’aigua necessàries, s’enfonsen en el mesofil·le o surten per la cara inferior de la fulla. Generalment a la fulla hi arriba una sola ramificació, que passa a través del pecíol. En arribar al limbe es torna a ramificar i origina una xarxa de nervadures vasculars que pot adoptar formes diferents.
El teixit lacunar és delimitat per sota per l’epidermis inferior. Les cèl·lules d’aquesta capa tenen bàsicament les mateixes característiques que les cèl·lules de l’epidermis superior, tret del gruix mínim de la cutícula, la presència ocasional de pèls, etc. Mentre que les cèl·lules de l’epidermis superior formen una capa contínua, les de l’epidermis inferior queden interrompudes aquí i allà per unes cèl·lules especials. Aquestes cèl·lules estan disposades per parelles formant obertures (els estomes) que comuniquen directament els espais intercel·lulars del teixit lacunar amb l’exterior.
Com que la cutina fa l’epidermis impermeable als gasos i l’aigua, és a través dels estomes que es produeixen els principals intercanvis gasosos entre la fulla i l’atmosfera. És, també, a través dels estomes que la planta pot regular la pèrdua d’aigua per transpiració.
Modificacions de les fulles
L’estructura foliar que hem descrit és la de la fulla "normal" (nomofil·le). Tanmateix, a la tija hi ha altres apèndixs laterals, també de tipus foliar, que es distingeixen del nomofil·le per l’aspecte, l’estructura, la posició que ocupen i la funció que realitzen. Aquests apèndixs es consideren modificacions del nomofil·le i per tant s’anomenen amb el terme genèric de "fulles modificades".
A més, en algunes plantes els nomofil·les, com a resultat de l’adaptació a determinades condicions i exigències ecològiques de la planta (clima àrid, nutrició carnívora, naturalesa enfiladissa, etc.), presenten un aspecte molt diferent del que hem descrit aquí. Aleshores es parla de metamorfosi de les fulles, i les estructures que en resulten s’anomenen "fulles metamorfosades". Volem remarcar que les flors també són modificacions de les fulles, però ja en parlarem amb més detall en el capítol 7.
Centrem-nos en les modificacions de les fulles. Les primeres fulles que es desenvolupen ja a l’interior de la llavor, als costats de l’àpex vegetatiu, són els cotilèdons. Les seves funcions es descriuen més endavant, als apartats "Fecundació i formació de l'embrió" i "La formació de la llavor".
Els hipsofil·les (o bràctees), en canvi, es troben en diverses posicions al llarg de les tiges. En el cas més típic se situen allà on un peduncle floral (o l’eix d’una inflorescència) s’insereix en la tija, i aleshores s’anomenen fulles axil·lars (perquè les flors neixen a la seva axil·la). També es poden trobar bràctees al llarg dels peduncles florals i, a vegades, immediatament sota la flor (o la inflorescència, en el cas que aquesta sigui una poncella). Constitueixen els anomenats sèpals i, contribueixen a la protecció de la flor. Els hipsofil·les en general presenten un desenvolupament reduït, un color entre marronós i verdós, i la clorofil·la hi és escassa o absent. Gairebé mai no tenen pecíol. Però de vegades assumeixen la funció de pètal superior pròpia de la corol·la, en les espècies en què és reduïda i poc vistosa. És el cas, per exemple, de la buguenvíl·lia, d’algunes eufòrbies, etc. En aquests casos les bràctees dels sèpals tenen colors molt vius i un desenvolupament igual o una mica inferior al dels nomofil·les.
Els catafil·les (o esquames) són a les tiges subterrànies. Embolcallen la gemma en els períodes en què cessa l’activitat vegetativa (per exemple durant l’hivern) i protegeixen els delicats teixits meristemàtics del fred, la penetració de l’aigua i la transpiració excessiva. Per això cobreixen ben bé la gemma i tenen un aspecte més aviat rugós i cuirassat. Moltes vegades són revestits de substàncies impermeabilitzants o d’una pelussa espessa, com en el salze. Sovint hi ha altres esquames que protegeixen els òrgans subterranis de la tija com els rizomes i els bulbs.
Com ja s’ha dit, els nomofil·les (les fulles "normals") poden presentar adaptacions morfològiques particulars en funció de la planta a la qual pertanyen. Vegem quines són les metamorfosis principals.
Les fulles suculentes són pròpies de les plantes que viuen en climes secs com el mediterrani (l’atzavara, l’àloe, les crassulàcies) o en sòls que no permeten un subministrament abundant i constant d’aigua (les plantes d’alta muntanya com les semprevives i les saxífragues, etc.). També són típiques de les plantes que viuen a les platges i als litorals on els sòls són fortament salins (la barrella). En aquest cas la fulla adquireix, a més de la funció assimiladora, la de dipòsit d’aigua o bé d’estructura segrestadora de sodi (cas de les plantes suculentes dels llocs salats, els halòfits).
La transformació de les fulles en espines és la metamorfosi més estesa i es dóna en plantes molt diferents. Generalment s’interpreta com una adaptació de la planta per a protegir-se de la mossegada dels herbívors. A vegades algunes fulles es transformen completament en espines (l’arç blanc, el roser, la robínia, el coralet), en alguns casos el limbe es desenvolupa normalment però el seu marge es torna espinós (el grèvol, el card, etc.), i en altres casos la punta de la fulla es transforma en espina (l’àloe, l’atzavara i la iuca). Hi ha algunes plantes, com la figuera de moro, en què totes les fulles s’han transformat completament en espines. L’explicació rau en el fet que en aquestes plantes la tija també pateix una metamorfosi per a esdevenir suculenta, tornant-se verda i assumir, per tant, la funció assimiladora.
Els circells són presents en algunes plantes enfiladisses, que per mitjà d’aquesta metamorfosi de les fulles s’agafen al suport on creixen. Els circells poden provenir de la transformació de tota la fulla o de només una part (vegeu també "Les plantes del nostre entorn").
En les plantes carnívores les fulles pateixen modificacions molt profundes que els permeten capturar els animals (insectes i altres invertebrats petits), dels quals extreuen la nutrició mineral (vegeu "Les fulles transformades de les plantes carnívores").
Les fulles transformades de les plantes carnívores
Entre les nombrosíssimes espècies de plantes, n’hi ha algunes que tenen la capacitat de nodrir-se de petits animals (com artròpodes, nematodes, petits crustacis). Després de capturar els animals, aquests són digerits per mitjà de sucs produïts per les mateixes plantes o amb l’ajut de bacteris o altres microorganismes simbionts. A la pràctica, aquestes plantes busquen un complement a l’alimentació mineral del sòl digerint material orgànic. Els mètodes adoptats per a la captura dels animals són diversos i molt interessants. Algunes espècies de Drosera, per exemple, tenen la superfície de les fulles plena de vellositats mòbils, que en l’àpex secreten un suc viscós. Quan l’insecte es col·loca sobre les fulles, és embolicat immediatament per les vellositats per tal d’atrapar-lo i després digerir-lo. Altres espècies, com les del gènere Nepenthes, natives de les zones humides tropicals del Vell Món, porten a l’àpex de les fulles un ascidi dotat d’una caputxeta mòbil que es tanca de cop per atrapar la presa. A l’interior de l’ascidi es distingeixen tres zones: la més propera a l’obertura és recoberta de glàndules secretores de nèctar, que serveix de reclam per als insectes; la central és formada per pèls dirigits cap avall que impedeixen a la presa retrocedir cap amunt, i la zona basal és recoberta de glàndules digestives. Es pot intuir fàcilment el destí d’un insecte que s’acosti a aquesta trampa.
Val la pena remarcar el fet que, llevat del Drosophyllum lusitanicum endèmic que viu als turons àrids de Portugal i el Marroc, totes les espècies carnívores són característiques de zones molt humides, lacustres, pantanoses, amb substrat tant sorrenc com fangós, on evidentment hi ha força aigua, però escassesa de nitrogen i sals minerals assimilables. Les plantes, per tant, han de "completar" la seva dieta i obtenir a partir d’organismes animals les substàncies que els falten.
La flor
La flor sol ser la part més vistosa de la planta, i és on tenen lloc els processos reproductius. És un òrgan característic de les angiospermes, mentre que en les gimnospermes té una estructura menys diferenciada i també es coneix amb el nom d’estròbil o con. A part de la seva importància funcional, pel que fa a la reproducció del món vegetal, la flor presenta una forma i una varietat de colors que la fan l’element més admirat de la natura.
Anatomia de la flor
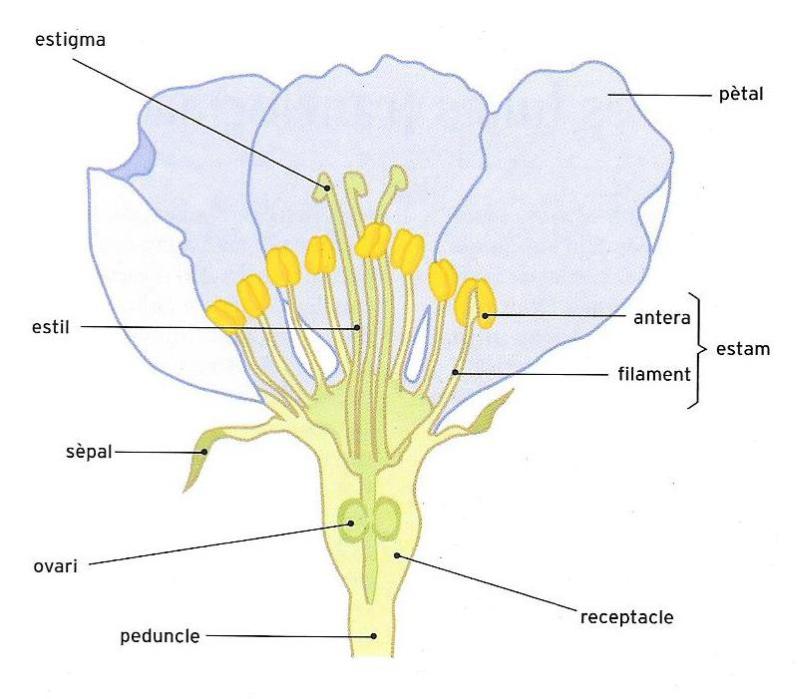
Estructura de la flor de les angiospermes, amb les parts externes visibles: el peduncle, el calze -format pels sèpals- i la corol·la -formada pels pètals-; i les parts internes: el gineceu -format per l'ovari, l'estil i l'estigma- i l'androceu -constituït pels estams.
ECSA
Les diverses parts de què consta la flor són formades, com ja s’ha dit, de fulles modificades. Algunes d’aquestes fulles (les antofil·les) constitueixen l’involucre floral o periant, i d’altres (les esporofil·les) l’aparell reproductor pròpiament dit. La modificació d’aquestes darreres és tal, que el seu aspecte no té res a veure amb una fulla.
En general, la flor se situa al capdamunt d’una estela o peduncle floral que acaba en un receptacle sobre el qual es disposen els diversos òrgans florals. El calze és la part més exterior de l’involucre floral i és constituït per diversos elements anomenats sèpals, que poden ser independents o estar units. Els sèpals solen ser poc espectaculars, de color verd i d’aspecte membranós o foliaci.
La corol·la, que juntament amb el calze forma el periant, és constituïda per diverses parts, també independents o bé unides, que s’anomenen pètals. Els pètals són generalment més grans que els sèpals i solen tenir colors vius (blaus, grocs, porpra, a vegades bigarrats, sovint blancs). La corol·la és la part més atractiva de la flor, és a dir, la part que atreu els insectes pol·linitzadors. Al seu interior, generalment a la base dels pètals, sovint hi ha glàndules nectaríferes, constituïdes per cèl·lules que elaboren i secreten el nèctar. El nèctar és un líquid "ensucrat" que agrada molt als insectes pol·linitzadors, els quals s’introdueixen en la flor per poder-ne extreure el pol·len.
A l’interior de la corol·la hi ha els òrgans reproductors pròpiament dits: l’androceu i el gineceu. L’androceu, o aparell masculí, és constituït per un nombre variable d’estams (que oscil·la entre dos i diverses desenes, segons les espècies). Els estams consten d’una part allargada, el filament, al capdamunt del qual hi ha una part més ampla, l’antera. L’antera se subdivideix en diversos sacs pol·línics, on es diferencia el teixit fèrtil de les cèl·lules que originen les cèl·lules germinals masculines (els gàmetes). En les plantes aquestes cèl·lules germinals masculines són els grans de pol·len. En moltes espècies els grans de pol·len són recoberts d’involucres molt resistents, capaços de romandre inalterats durant mil·lennis. Precisament a partir de l’anàlisi del pol·len fòssil dipositat al sòl podem obtenir molta informació sobre les plantes d’èpoques anteriors.
En madurar, els teixits externs de l’antera s’esquerden i els grans de pol·len madurs són alliberats a l’exterior perquè puguin dur a terme la seva funció.
A l’interior de la corona d’estams, a la part més central i més profunda de la flor, hi ha l’aparell reproductor femení o gineceu. És constituït per una o més fulles anomenades fulles carpel·lars o carpels que, en replegar-se, formen una cavitat tancada anomenada ovari. Dins l’ovari hi ha els òvuls, en els quals es diferencien les cèl·lules germinals femenines (els gàmetes). Un cop fecundats pels grans de pol·len, els òvuls es modifiquen i es desenvolupen per a originar la llavor. El gineceu pot ser constituït d’un sol carpel, que, en replegar-se sobre ell mateix, origina un ovari unicarpel·lar, o de diversos carpels, que creixen i s’uneixen entre ells formant un ovari pluricarpel·lar.
La forma més típica del gineceu és constituïda per una base arrodonida i completament buida, l’ovari, coronada per una prolongació anomenada estil, que al capdamunt tendeix a eixamplar-se una mica per a formar l’estigma.
L’estigma representa, per dir-ho d’alguna manera, el "punt d’acollida" dels grans de pol·len que, transportats pel vent o pels animals, comencen el camí que els portarà fins al fons de l’ovari. El conjunt constituït pel gineceu, l’estil i l’estigma s’anomena pistil.
Diversos tipus de flors i inflorescències
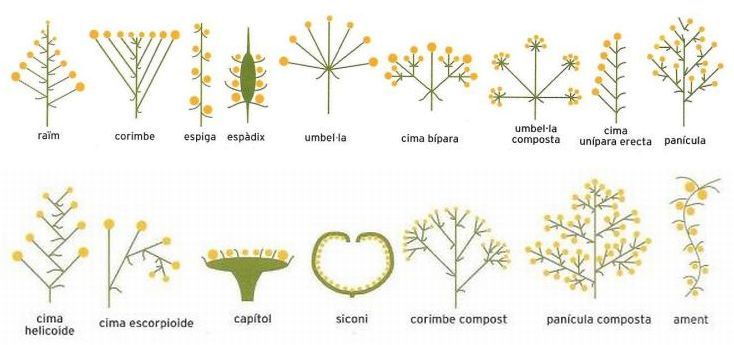
Esquema de diferents tipus d'inflorescències. Hi podem reconèixer les espigues de blat, els paraigües (umbel·la) de la pastanaga bord, la forma dels raïms, els capítols de les margarides i, fins i tot, el cas especial del siconi de les figues.
ECSA
Les parts que constitueixen la flor s’insereixen en el receptacle seguint les mateixes modalitats que regulen la inserció de les fulles al brot. Poden disposar-se aïlladament al llarg d’una espiral (flors acícliques) o en verticil (flors cícliques), o bé d’una manera combinada (flors hemicícliques). El nombre de pistils, d’estams i d’antofil·les, la seva disposició en el receptacle i la seva conformació són molt importants des d’un punt de vista sistemàtic i s’utilitzen per a la caracterització i la identificació de les diverses famílies, gèneres i espècies.
A més, segons la conformació del receptacle es distingeix entre flors hipògines o flors amb ovari súper, flors perígines i flors epígines o amb ovari ínfer. En les flors hipògines, el receptacle té forma de disc i el pistil inserit al centre es troba per damunt de tots els altres elements florals. En les flors perígines el receptacle té forma de copa, el pistil s’insereix al fons al centre, i els estams i els antofil·les, a la vora. En les flors epígines, la part inferior del pistil, l’ovari, s’inclou en el receptacle i, per tant, queda una mica més avall del punt d’inserció de la resta d’elements florals. A més, els elements de la flor poden ser lliures, és a dir, independents els uns dels altres (corol·les dialipètales), o estar units (corol·les gamopètales o simpètales). Les corol·les amb els pètals units poden tenir una conformació molt diferent, segons si els pètals s’uneixen entre ells completament o només en part. En el primer cas prevalen en general les formes d’embut o de didal, mentre que en el segon la part inferior de la corol·la sol ser tubular o s’obre cap amunt, presentant lòbuls separats, més o menys desenvolupats, i amb orientacions diverses.
També és important la simetria floral, que pot ser radiada (flors actinomorfes o en estrella) o bilateral (flors zigomorfes). En la majoria de les angiospermes, la corol·la cau després de la pol·linització, perquè ja ha realitzat la seva funció primordial d’atreure els insectes pol·linitzadors. En canvi, el calze sol persistir i inicialment protegeix el pistil que s’està transformant en fruit.
Les flors poden ser aïllades, formar-se a l’extrem dels brots o a l’axil·la de les fulles, o bé es poden agrupar en inflorescències. Entre els tipus més comuns d’inflorescència trobem: el raïm simple, amb flors pedunculades; l’espiga, amb flors sèssils; el corimbe, que té els peduncles de les flors de llargades diferents i aleshores totes queden al mateix nivell; el capítol, amb petites flors sèssils inserides a l’àpex de l’eix floral, engruixit per formar un receptacle; la umbel·la, d’aspecte semblant al corimbe, però en què tots els peduncles van a parar a l’extrem de l’eix. També són freqüents les inflorescències compostes, com la umbel·la composta, la panícula (o raïm compost), el corimbe compost i d’altres. Finalment, hi ha les inflorescències cimoses, com la cima unípara i la cima bípara.
Les flors i les inflorescències s’associen sovint a fulles modificades quant al color o la forma, com és el cas de l’eufòrbia, que té les fulles que acompanyen les inflorescències d’un color vermell molt viu, o bé constituint bràctees amples com és el cas de l’Anthurium o la cal·la. En aquests casos, o bé les fulles substitueixen la corol·la pel que fa a la funció d’atreure els insectes perquè pol·linitzin, o bé serveixen de protecció.
Llavor i fruit: la reproducció de les plantes superiors
En la majoria dels casos les plantes es reprodueixen per gamogènesi, és a dir, sexualment. Neixen i es desenvolupen d’una llavor, sorgida de la unió d’un gàmeta femení i un de masculí (fecundació). Però també és molt estesa la reproducció asexual (anomenada propagació o multiplicació vegetativa), que comporta la formació de nous individus a partir de cèl·lules o parts de la planta mare. Un tercer tipus de reproducció, també important, és l’esporògenes, que té lloc gràcies a cèl·lules especials, les meiòspores, dins un cicle que també inclou la producció de gàmetes. La multiplicació vegetativa i l’esporogènesi seran tractades més breument en els inserts.
Comencem, doncs, per la llavor: de què es tracta, concretament? Com es forma? Per a respondre la primera pregunta podem dir que la llavor és una mena de planteta en miniatura que duu una vida alentida i, per tant, amb necessitats molt reduïdes. La llavor pot sobreviure durant molt temps, fins i tot en condicions ambientals adverses. Pot, per tant, esperar el retorn de les condicions favorables, i quan arribin, podrà germinar i produir una nova planta.
Els mecanismes de la reproducció
La reproducció dels vegetals superiors no és, des del punt de vista genètic, diferent de la dels animals. En les plantes el patrimoni genètic també és contingut en els cromosomes, el nombre dels quals és característic i constant en cada espècie. Per tal que aquest nombre no es dupliqui a cada generació, les cèl·lules germinals masculines i femenines (els gàmetes) que s’uneixen per a formar el zigot, del qual naixerà la llavor i la nova planta, tenen la meitat de la dotació cromosòmica (és a dir, són cèl·lules haploides). Aquestes cèl·lules provenen d’una cèl·lula mare que ha patit una divisió reductora, és a dir, la meiosi. Quan les dues cèl·lules germinals amb la meitat de dotació s’uneixen per a formar el zigot, es recupera el nombre cromosòmic característic de l’espècie i el patrimoni genètic essencial perquè la planta pugui assolir el desenvolupament complet i acomplir correctament les seves funcions vitals.
Però, si bé la reproducció dels vegetals és perfectament anàloga a la dels animals quant a la genètica, no es pot dir el mateix de les modalitats d’unió dels dos gàmetes. Les plantes, organismes fixats al sòl, no tenen, evidentment, la possibilitat d’apariar-se, i d’altra banda els seus òrgans reproductors no estan preparats per a fer-ho. La majoria de les plantes tenen flors de tots dos sexes. Així, podríem pensar que superen l’obstacle que representa la seva immobilitat autofecundant-se. Perquè, què seria més fàcil, còmode i segur per a una planta que fer caure els grans de pol·len alliberats per les anteres a l’ovari de la mateixa flor, i produir així un zigot (a partir del qual un embrió i una llavor) capaç de reproduir-la? Però si això fos així la planta no podria gaudir dels avantatges d’encreuar-se amb altres individus de la mateixa espècie.
En efecte, l’experiència demostra que l’autofecundació no tan sols implica l’acumulació progressiva de tares genètiques que empitjoren de generació en generació les característiques i la capacitat de supervivència dels descendents obtinguts així, sinó que també, i sobretot, exclou la barreja dels gens, font de la variació indispensable perquè l’espècie es pugui adaptar a les condicions ambientals canviants. Per evitar l’autofecundació i els riscs que se’n deriven, les plantes disposen d’un recurs específic: que els òrgans masculins i femenins madurin en temps diferents. Quan els sacs pol·línics d’una flor maduren i alliberen el pol·len, l’aparell femení encara és immadur o, al contrari, ha madurat abans i ja ha estat fecundat pel pol·len d’un altre individu. Així, els microscòpics grans de pol·len han de viatjar cap a una flor d’un altre individu de la mateixa espècie.
L’alternança de generacions en les plantes
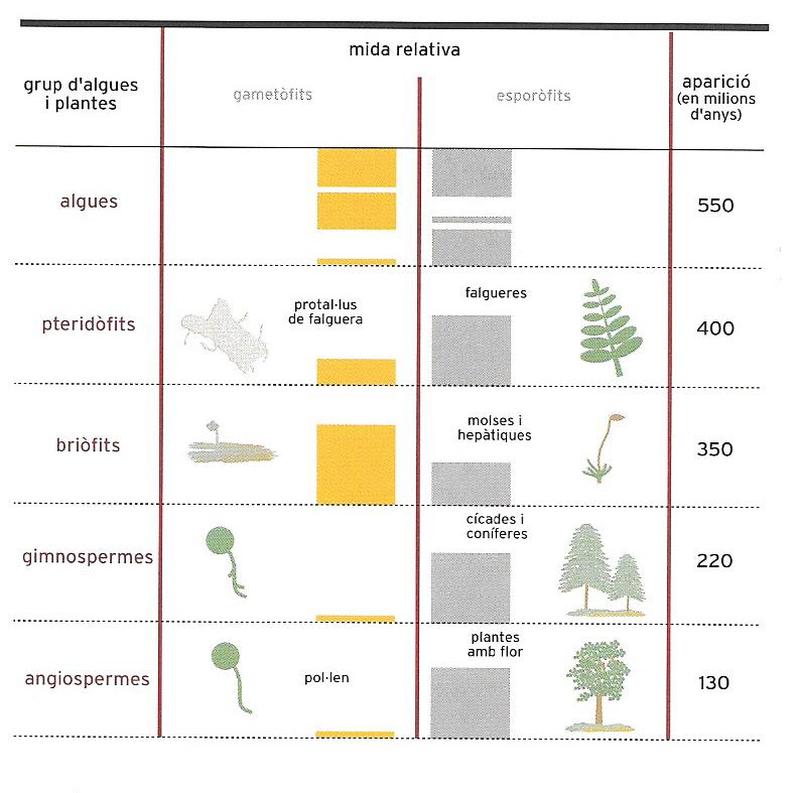
Alternança de generacions als principals grups d'algues i plantes. Al llarg de l'evolució, la importància relativa de l'esporòfit sobre el gametòfit com a generació dominant de la forma vegetativa ha anat augmentant. En els algues alternen les fases haploide -representada pel gametòfit- i diploide -representada per l'esporòfit-, tant si el gametòfit i l'esporòfit són iguals (cicle isomòrfic), com si són diferents (cicle heteromòrfic), en aquest últim cas, el gametòfit o l'esporòfit poden ser la generació dominant de la forma vegetativa. En els briòfits la situació és bastant diferent, ja que l'esporòfit generalment és més petit i viu sobre el gametòfit depenent d'aquest. En els pteridòfits ja domina clarament la generació esporofítica, bé que els gametòfits tenen una existència independent. La màxima reducció dels gametòfits es dóna en les gimnospermes i angiospermes, on són microscòpics i depenen dels esporòfits.
ECSA
En les plantes existeixen tres tipus de cicles: l’haploide, l’haplodiplont i el diploide. En els casos típics es tracta d’una alternança regular de dues generacions representades per individus que morfològicament sovint són diferents i que produeixen cèl·lules germinals distintes. L’esporòfit, o generació diploide, produeix espores mitjançant meiosi i el gametòfit, o generació haploide, produeix gàmetes per mitosi.
La condició primitiva és l’haploide, o sigui, organismes que amb una única dotació cromosòmica (n) es multipliquen asexualment. En algun moment es produí la fusió de dos organismes haploides i s’originà un organisme diploide, amb dotació cromosòmica (2n), que, almenys al principi, per tal de ser viable hauria de patir de manera immediata una meiosi que el retornés al seu estat haploide. En aquest moment, amb la fusió de nuclis apareix la sexualitat i alternança de fases nuclears, haploide/diploide, i es multiplica el nombre de nous genotips i, per tant, el material de selecció. Aquest cicle, típicament haplont i dominat pel gametòfit, és el que representen algues primitives i moltes algues verdes. Posteriorment, la meiosi s’anà retardant fins a produir-se en l’esporòfit desenvolupat a partir del zigot. De forma simultània la condició diploide ha estat seleccionada positivament, ja que un organisme amb dotació diploide té un joc genètic complet de reserva que accepta mutacions letals en recessivitat.
Els talassòfits, algues bentòniques que són representants vegetals característics del Cambrià, Ordovicià i Silurià, ja tenien alternança de generacions isomòrfica, és a dir, amb el gametòfit i l’esporòfit morfològicament iguals. Aquest tipus de cicle és molt comú en altres tipus d’algues, com per exemple en Ulva. Altres, com Nereocystis, tenen el gametòfit molt reduït.
Les plantes terrestres primitives tenien un cicle isomòrfic que es va anar transformant en heteromòrfic amb predomini de l’esporòfit. No obstant això, els briòfits s’anaren adaptant a les condicions del medi terrestre basant-se en el gametòfit i desenvolupant-hi a sobre l’esporòfit. Les plantes que demostraren ser més adequades per a la conquesta de la terra foren els pteridòfits, que prèviament havien desenvolupat una generació esporògena diploide resistent, ja que aconseguí impedir millor la dessecació. En els pteridòfits, el gametòfit és força reduït, però a diferència dels briòfits no està lligat a l’esporòfit. L’aparició de l’heteroscòpia en els pteridòfits (fa uns 360 milions d’anys) fou molt important perquè el seu èxit en algunes línies d’aquest grup conduí a l’origen dels espermatòfits, que també són heterospòrics. Les espècies amb heterospòria produeixen micròspores i megàspores en els gametòfits masculí i femení, respectivament, assegurant-se d’aquesta manera la separació de sexes. Els equisets tenen cicle intermedi entre isospòric i heterospòric amb heterospòria funcional.
En les fanerògames, malgrat que el gametòfit s’ha reduït a un conjunt de poques cèl·lules, el masculí és encara independent, però resta limitat a l’interior del gra de pol·len; el femení viu com a paràsit de l’esporòfit inclòs dins els teixits de l’òrgan femení de la flor. La gran innovació dels espermatòfits és la dispersió mitjançant llavors. Això, juntament amb la seva adaptació al medi terrestre, ha provocat l’aparició d’una o més capes protectores de les megàspores, una en el cas de les gimnospermes i dues en el cas de les angiospermes.
En les gimnospermes es produeixen dues especialitzacions: per una banda es continuen reduint els gametòfits i per l’altra es passa de la zoidogàmia —fecundació mitjançant cèl·lules flagel·lades— en les primitives, com Cycas i Ginkgo, a la sifonogàmia —fecundació a través del tub pol·línic amb cèl·lules no flagel·lades— en les més evolucionades, com Pinus. Els estams de les angiospermes contenen 4 sacs pol·línics o microsporangis, mentre que en les gimnospermes el nombre és molt variable.
En les angiospermes trobem la màxima reducció del gametòfit, el qual la seva vida lliure, però resta protegit de la dessecació en el medi terrestre. El femení, en la seva maduresa únicament està format per 8 cèl·lules. L’altra novetat és la doble fecundació on, per una banda, s’origina l’embrió i, per l’altra, l’endoderma. El masculí, extremament reduït, està format exclusivament per tres cèl·lules. Prova de l’èxit evolutiu d’aquest tipus de cicle és que en l’actualitat existeixen unes 260 000 espècies d’angiospermes mentre que tan sols n’hi ha 10 800 de la resta d’espècies terrestres.
La importància ecològica de l’alternança de generacions rau en l’especialització d’aquestes. Així, la generació asexuada, l’esporòfit, assegura un nombre alt d’individus en poc temps, com és el cas de moltes espècies de plantes superiors que es multipliquen vegetativament; però el gametòfit, la generació sexuada, recombina de tant en tant el material genètic.
La pol·linització
Els grans de pol·len viatgen d’una flor a l’altra sobretot de dues maneres: per mitjà del vent o per mitjà dels animals, gairebé sempre insectes. En el primer cas es parla de pol·linització anemòfila, i en el segon de pol·linització zoòfila (del grec "anemos", vent, i "zoon", animal, respectivament).
Pel que fa a la pol·linització provocada pel vent, semblaria que el fet que els grans de pol·len aconsegueixin o no el seu destí depèn exclusivament dels capricis incontrolables dels corrents d’aire i, en definitiva, de l’atzar. I certament és així, però, a causa del gran nombre de grans produïts (una sola antera pot produir molts milions d’aquestes partícules microscòpiques), per una simple qüestió de probabilitat estadística és gairebé segur que alguns grans arribaran a bon port, és a dir, al cim de l’ovari (l’estigma) d’una flor de la mateixa espècie. A més, com que per a fecundar un aparell femení n’hi ha prou amb un sol gra de pol·len, encara que la quantitat de grans que hi arriben sigui petita és suficient per a fecundar moltes flors i, per tant, per a garantir la reproducció de l’espècie.
Amb la pol·linització zoòfila sembla que el destí de les espècies depèn d’un procés més segur i menys casual. Els pol·linitzadors més comuns són els insectes, però també molts altres animals, com els ocells (els colibrís, per exemple) i petits mamífers (alguns ratpenats, rosegadors i marsupials), són distribuïdors actius del pol·len. Els insectes visiten les flors amb l’objectiu de xuclar el nèctar contingut a les glàndules que generalment hi ha a la base de la corol·la. Per a arribar a aquestes glàndules els insectes han de penetrar pràcticament fins al fons de la flor, amb la qual cosa sacsegen les anteres i provoquen una pluja de pol·len que els va a parar precisament al damunt del cos i les potes.
Un cop ha obtingut el nèctar, l’insecte continua passejant-se entre les flors bo i transportant el pol·len de les diferents flors als ovaris de moltes altres. Aquesta acció inconscient és beneficiosa tant per a la planta que s’ha pol·linitzat, com per a l’insecte perquè d’aquesta manera també afavoreix el creixement d’un gran nombre de plantes que són la seva font d’aliment.
És sorprenent la perfecta complementarietat entre les característiques de la flor i les de l’insecte pol·linitzador. També sorprenen els recursos mostrats per les plantes per a atreure els insectes. Per exemple, la forma i les dimensions de la corol·la, la posició dels estams i de l’ovari poden forçar l’insecte a fer un recorregut i uns moviments concrets i, per tant, a adoptar postures obligades dins la corol·la que tant poden afavorir que l’insecte perdi el pol·len recollit d’altres flors com que "recarregui" pol·len alliberat per les anteres d’aquesta flor.
Les flors, a més, poden emetre olors similars a les de les femelles de les espècies pol·linitzadores, i així atreuen els mascles corresponents. Altres vegades l’aspecte de la flor recorda les femelles de determinats insectes, com és el cas del gènere Ophrys de la família de les orquídies.
Les adaptacions recíproques entre la planta i el pol·linitzador arriben a una situació extrema en els casos en què una planta depèn d’una sola espècie d’insecte per a ser pol·linitzada. Quan els insectes responsables de la pol·linització pertanyen a més d’una espècie, les adaptacions són menys notables. Amb tot, a grans trets, les plantes de pol·linització zoòfila tenen flors grans, de colors vius, capaces, per un o altre motiu, d’atreure i acollir al seu interior l’animal o els animals pol·linitzadors. Entre aquestes plantes poden destacar les de la família de les rosàcies (a la qual pertanyen els rosers i els principals arbres fruiters), les orquídies, els lliris, els ranuncles i el romaní.
En les plantes de pol·linització anemòfila (és a dir, per mitjà del vent), les flors tendeixen a ser petites, tenen un color semblant al de la resta de la planta i en conjunt són poc vistoses. Les corol·les grans no solament serien inútils, perquè no cal cap reclam visual, sinó també un obstacle per a escampar el pol·len. En posarem alguns exemples. La majoria dels arbres (els faigs, els bedolls, els carpins, els castanyers, les alzines, les nogueres i totes les coníferes) són de pol·linització anemòfila. Entre les plantes herbàcies cal destacar les gramínies (el blat, l’arròs, el blat de moro, etc.).
La formació de l'òvul
El gra de pol·len portat pel vent o els animals arriba, doncs, al seu destí, és a dir, a l’àpex de l’ovari que conté els òvuls. La quantitat i la disposició dels òvuls és diferent en les diverses espècies.
A l’interior de l’òvul hi ha un teixit anomenat nucel·la i dos teguments al voltant que no acaben d’embolcallar l’òvul del tot, sinó que deixen una obertura anomenada micròpil. I és a través d’aquesta obertura que hi penetra el gra de pol·len. A la part més profunda de la nucel·la es produeix la divisió per meiosi, que origina cèl·lules haploides. La majoria d’aquestes cèl·lules degeneren i moren, però n’hi ha una, i només una, que creix notablement, forma grans vacúols i origina una estructura anomenada sac embrional que se situa a l’interior de la nucel·la.
El nucli d’aquesta cèl·lula es torna a dividir i origina vuit nuclis més (també haploides), dels quals tan sols n’hi ha un destinat a ser fecundat pel gra de pol·len i que origina l’embrió. Aquest nucli és el veritable gàmeta femení i rep el nom d’ovocèl·lula.
Fecundació i formació de l'embrió
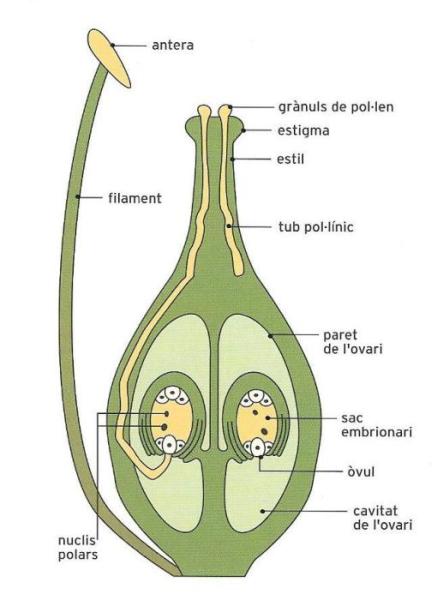
Esquema simplificat de la fecundació. Just abans d'iniciar-se, dins el gra de pol·len, s'hi han diferenciat una cèl·lula vegetativa i dues d'espermàtiques. Dins l'òvul s'han format vuit nuclis, dels quals únicament l'ovocèl·lula està destinada a contribuir a la formació del zigot. Es diu que la fecundació és doble perquè, mentre un dels nuclis espermàtics es fusiona amb l'ovocèl·lula, l'altre ho fa amb els dos nuclis polars per a formar l'anomenat endosperma, que serà la substància de reserva de la llavor.
ECSA
Hem deixat el gra de pol·len dipositat sobre l’estigma. A partir d’aquí el gra fa l’última part del recorregut cap a l’ovocèl·lula emetent una protuberància o filament prim que s’anomena tub pol·línic, cap a l’interior del qual migra el nucli (haploide) del gra.
En allargar-se, el tub penetra en l’ovari. Quan ateny l’extrem del sac embrional, on hi ha l’ovocèl·lula, el nucli pol·línic i el de l’ovocèl·lula es poden fondre i donar origen a una cèl·lula que torna a tenir dotació genètica diploide, el zigot.
A partir de les divisions del zigot es forma el primer esbós de la planta: l’embrió. La llavor és el producte de la maduració de l’òvul després d’haver estat fecundat. Hi ha un moment en què l’embrió es diferencia morfològicament, de manera que s’hi distingeixen tres parts: la radícula, la plúmula i els cotilèdons. La radícula és la part d’on creixerà l’arrel quan la llavor germini. La plúmula constitueix l’àpex de l’embrió, a partir del qual s’originarà la tija amb les seves ramificacions i els seus apèndixs. És, doncs, tota la part no subterrània de la planta. Els cotilèdons, que poden ser dos o un de sol (depenent d’això les angiospermes se subdivideixen en dos grans grups: les dicotiledònies i les monocotiledònies), es desenvolupen i tenen unes funcions variades segons les diverses espècies. Mentre l’embrió es desenvolupa fins a originar l’estructura que hem descrit, a la resta de l’òvul es donen tot un seguit de processos de maduració que finalitzen en la transformació de l’òvul en llavor.
La formació de la llavor
Ni l’embrió ni la planteta que germinarà (la plàntula) tenen la capacitat d’autosostenir-se, perquè no posseeixen teixits dotats d’activitat fotosintètica i encara no tenen un sistema radical que els permeti absorbir les sals minerals del sòl. La planta mare, que acumula les substàncies nutritives en un teixit especial del sac embrional (l’endosperma secundari) o en altres parts de l’òvul o de l’embrió, s’encarrega de nodrir-lo.
El pas de l’òvul a llavor pot seguir modalitats diferents, segons el teixit en què es dipositen les substàncies de reserva. Podem distingir:
1) Les llavors que concentren les seves reserves en l’endosperma. En aquest cas l’endosperma presenta un desenvolupament notable: l’embrió s’enfonsa al seu interior, i la nucel·la desapareix del tot o es redueix a un prim involucre que cobreix l’endosperma. Són llavors d’aquesta mena, per exemple, les de les gramínies, molt importants des del punt de vista alimentari. El pa, la pasta i tots els productes a base de midó procedeixen de l’endosperma de les llavors de les gramínies, i el seu valor alimentari és conegut pels humans des de fa molt temps.
2) Les llavors que concentren les seves reserves en els cotilèdons. Els cotilèdons assoleixen, aleshores, un desenvolupament molt notable, mentre que l’endosperma pràcticament no es forma i la nucel·la desapareix o es redueix molt. Una altra gran família de plantes, també molt important per a l’alimentació humana, que ens pot servir d’exemple són les lleguminoses (família a la qual pertanyen el pèsol, la mongeta, la llentia, el cigró, la soja, etc.). Quan mengem un d’aquests llegums, allò que ens alimenta són precisament les seves reserves, que contenen molt de midó i proteïnes. Una llavor de mongetera ens mostra d’una manera molt evident quina és la part del volum que s’ha d’atribuir als cotilèdons carnosos i engrandits, entre els quals el petit embrió queda comprimit.
3) Les llavors la funció de reserva de les quals és assumida per la nucel·la. Aleshores la nucel·la no pateix cap reducció a costa de l’endosperma, mentre que aquest presenta un desenvolupament escàs o nul i els cotilèdons un desenvolupament normal. Són d’aquest tipus, per exemple, les llavors de la planta del pebrer.
Per a completar l’estructura de la llavor encara falten els dos teguments de l’òvul, que durant els processos de maduració es modifiquen per tal de poder assumir amb eficàcia la funció de protecció de l’embrió. Esdevenen coriacis i impermeables per a poder oposar resistència tant als eventuals organismes portadors de malalties (els fongs, els bacteris, etc.) com a la penetració d’aigua. La hidratació, és a dir, que la llavor s’impregni d’aigua és una condició essencial perquè germini. Però moltes llavors cauen a terra en un moment en què la germinació no és oportuna; aleshores, els teguments han d’endarrerir-la bo i impedint que l’aigua penetri dins la llavor. Penseu, per exemple, en una llavor produïda a la tardor, cosa habitual en moltes espècies. Si la llavor germinés de seguida, ben aviat la plàntula estaria exposada al rigor de l’hivern.
El fruit
Després de la fecundació, mentre es produeix en l’òvul la transformació de l’embrió en llavor, l’ovari també pateix profundes modificacions que el porten a convertir-se en fruit. Tots sabem, o creiem que sabem, què és un fruit. Una poma, una pera, una cirera, un préssec o una taronja són fruits; però un tomàquet, un pebrot i una mongeta també ho són. Totes les plantes produeixen fruits. En alguns casos, com els que hem esmentat, aquests fruits tenen característiques que fan que per a nosaltres, els humans, siguin comestibles; en d’altres, els fruits poden representar un recurs alimentari important per als animals salvatges. Però hi ha fruits que no són comestibles ni per als humans ni per als animals, i es limiten a acomplir la funció de "receptacles" temporals de les llavors.
Normalment, el fruit prové directament del desenvolupament de la paret de l’ovari transformada en pericarp, que conté les llavors al seu interior. En alguns casos les transformacions necessàries per a la formació del pericarp tan sols afecten les dimensions i la consistència de la paret de l’ovari, mentre que altres vegades se’n modifiquen el volum, el gruix, la consistència, el color, la composició química i la forma. Aquests fenòmens tan variats tenen com a conseqüència una varietat extrema de fruits. La paret de l’ovari, que sol ser constituïda per un nombre relativament limitat de capes de cèl·lules, s’engruixeix a mesura que avança la formació del fruit. Quan és complet es distingeixen dues capes, una d’externa i una d’interna. L’externa, anomenada epicarp, inclou l’epidermis i les capes cel·lulars de sota. Per fora pot ser llisa i coberta d’una capa cerosa o acuminada. La capa interna, anomenada endocarp, sovint és lignificada i conté les llavors. En molts casos es forma, a més, una tercera capa intermèdia, anomenada mesocarp, carnosa, fibrosa i de consistència diversa. Així, per exemple, en la cirera, l’epicarp es la pell, el mesocarp correspon a la polpa i l’endocarp són les cèl·lules endurides i pètries que constitueixen el pinyol.
La gran varietat de fruits
La conformació dels fruits és molt variable i depèn principalment de la forma, de la constitució, de la posició en la flor i del nombre d’ovaris dels quals deriven. En els anomenats falsos fruits els teixits del receptacle també participen en la formació del fruit. Quan en una mateixa flor hi ha presents diversos ovaris, en resulten fruits compostos de molts fruits petits. Hi ha inflorescències compostes que poden formar fruits compostos anomenats infructescències. Els fruits es poden subdividir fonamentalment en fruits secs i fruits carnosos, segons si el pericarp és membranós o està format per teixits parcialment suculents.
Els fruits secs indehiscents són aquells fruits que quan maduren cauen de la planta sense obrir-se per a alliberar les llavors. Sovint tenen apèndixs alats o pilosos (vil·lans), que faciliten la seva difusió pel medi i per tant tenen un paper fonamental en la disseminació. Entre aquests fruits trobem els aquenis, característics, per exemple, de les diferents espècies de ranuncles i pixallits, en les quals és visible un vil·là molt desenvolupat: les sàmares alades de l’auró i el freixe. Les núcules, en canvi, tenen un pericarp llenyós molt resistent i generalment contenen una sola llavor. Sovint són envoltades per una cúpula que prové del receptacle o del peduncle floral, com la gla de l’alzina, o de les bràctees, com l’ametlla. La cariopsi de les gramínies és un cas concret on el pericarp del fruit i el tegument de la llavor formen un tot indivisible.
Els fruits secs dehiscents no cauen de la planta. Quan són madurs s’obren de diferents maneres per a alliberar les llavors, que gairebé sempre contenen en un nombre elevat. Recordem el llegum típic de les lleguminoses, que s’obre al llarg de la sutura ventral, i els diversos tipus de càpsules. Són característiques la càpsula de les roselles, que allibera les llavors a través d’una sèrie d’obertures disposades en cercle a la paret superior, i la càpsula del lliri, que s’obre al llarg de les línies verticals. El fruit de les crucíferes és una silíqua, amb un septe central on s’insereixen les llavors, que s’obre per la separació de dues valves laterals.
Els fruits carnosos tenen el pericarp totalment o parcialment carnós. La drupa té el pericarp carnós per fora i xilematós per dins, on hi ha el pinyol. Els animals que es nodreixen d’aquests fruits solen ser els encarregats de la seva disseminació. En són exemples típics la cirera, l’oliva i la nou (la part carnosa de la qual no és comestible). El coco també és una drupa, encara que una mica especial ja que el pericarp és estructurat de tal manera que pot surar i disseminar-se a través de la mar d’una illa a l’altra. La baia, en canvi, té tot el pericarp carnós, i normalment conté nombroses llavors revestides d’un tegument seminal resistent, com per exemple el tomàquet, el raïm, el meló i d’altres. Les baies normalment també són menjades pels animals que ingereixen les llavors, que, malgrat tot, no es veuen danyades pels processos digestius i, per tant, són disseminades amb les deposicions. En alguns casos, aquest pas a través del sistema digestiu dels animals és indispensable per a la germinació de les llavors. L’hesperidi del taronger és un tipus especial de baia.
L’exemple més típic de fals fruit és el pom. Són poms, per exemple, els fruits de la pomera, la perera i el codonyer, en els quals tota la part carnosa perifèrica prové del receptacle.
Els fruits compostos poden ser formats per moltes drupes petites ben apinyades, com el gerd o la móra, o per aquenis inserits en un receptacle carnós, com la maduixa, o de moltes altres maneres.
Les infructescències resulten d’inflorescències compactes, com per exemple els fruits de la morera, que són aquenis petits agrupats i units per teixits carnosos provinents del periant. Un cas del tot especial és la figa, en què tot l’eix floral esdevé carnós i presenta una cavitat interna on es formen moltes drupes petites.
Els teixits vegetals
També en les plantes, com en els animals, el conjunt de diverses cèl·lules amb una mateixa funció forma un teixit. Vegem quins són els principals teixits vegetals i quines són les seves característiques i funcions, partint de la subdivisió bàsica entre teixits meristemàtics o embrionaris i teixits adults. Tal com hem vist més amunt, els primers són constituïts per cèl·lules capaces de dividir-se, i els segons per cèl·lules plenament diferenciades que no tenen aquesta capacitat. A més, pel que fa als teixits adults, segons les diferents funcions que realitzen podem distingir: els tegumentaris, els parenquimàtics, els mecànics (o de sosteniment), els conductors i els secretors.
Els teixits meristemàtics
Com ja hem vist abans, les cèl·lules meristemàtiques són les encarregades de produir elements cel·lulars nous per al creixement de les plantes. Els teixits meristemàtics es divideixen en primaris i secundaris: els primers allarguen la planta i els segons l’engruixeixen. Els meristemes primaris es troben en l’àpex de les tiges i de les arrels. Les cèl·lules d’aquests meristemes tenen un aspecte "embrionari" típic, no diferenciat. Sempre provenen d’altres cèl·lules meristemàtiques, per més enrere que ens remuntem en el temps... fins arribar a l’òvul fecundat a partir del qual s’origina la planta sencera.
Els meristemes secundaris, en canvi, deriven de cèl·lules adultes que, com sempre, perden la capacitat de dividir-se durant la diferenciació, però la tornen a adquirir més tard. Els meristemes secundaris també es troben a les tiges i arrels, però no als àpexs tendres, sinó a la part de sota, on l’òrgan és massís i llenyós. En aquesta zona els meristemes secundaris formen dos anells concèntrics. Les cèl·lules d’aquests meristemes es divideixen preferentment en plans paral·lels a la superfície de l’òrgan, cosa que origina necessàriament un augment del gruix. Les cèl·lules dels meristemes secundaris no presenten el típic aspecte "de cèl·lula meristemàtica", sinó que conserven nombroses característiques de les cèl·lules adultes de les quals deriven, com per exemple els vacúols ben desenvolupats.
Els teixits tegumentaris
Els teixits tegumentaris revesteixen tota la planta exteriorment i tenen com a funció principal protegir-la contra una pèrdua excessiva d’aigua per evaporació. Això explica per què en tots els teixits tegumentaris les cèl·lules estan molt juntes, sense espais intercel·lulars. Les parets són parcialment constituïdes per cutina o suberina, que les fan impermeables a l’aigua.
El teixit tegumentari que revesteix les parts no llenyoses de les plantes (les tiges herbàcies, les fulles, les flors, els fruits...) rep el nom d’epidermis. Les cèl·lules epidèrmiques se solen disposar en un sol estrat: són vives, i tenen els vacúols grans i els plastidis poc desenvolupats. La grandària del vacúol les fa transparents a la llum, característica molt útil atès que l’epidermis sovint recobreix teixits fotosintètics. La cara externa de la paret cel·lular és impregnada de cutina. La impermeabilització augmenta gràcies a una capa contínua de cutina anomenada cutícula, que revesteix per fora com si es tractés d’un llençol totes les cèl·lules de l’epidermis. En alguns casos, per sobre de la cutina hi ha un segon revestiment cerós que augmenta encara més la impermeabilitat. És fàcil reconèixer un òrgan revestit de ceres (per exemple la pell del raïm, la fulla d’una planta crassa, la tija del blat) perquè si se submergeix en aigua no es mulla.
L’epidermis té unes obertures regulables —comparables a vàlvules— que s’anomenen estomes (del grec "stoma", boca). A cada mil·límetre quadrat d’epidermis s’obren desenes i sovint fins i tot centenars d’estomes. Són necessaris perquè la cutina, a més de ser impermeable a l’aigua, també ho és als gasos. Si no hi fossin, no solament es blocaria l’evaporació sinó també l’entrada del diòxid de carboni, amb la qual cosa seria impossible portar a terme l’activitat fotosintètica. A més, si les obertures fossin permanents, no regulables i per tant no es poguessin tancar en cas d’emergència hi podria haver el perill que la planta s’assequés. La regulació permet assolir l’obertura màxima compatible amb la disponibilitat d’aigua del moment.
Sovint, a l’epidermis també hi ha pèls que poden estar formats per cèl·lules vives o per cèl·lules mortes i que tenen funcions diverses (la secreció, l’impedir l’evaporació i l’escalfament excessius, etc.).
Un altre teixit tegumentari és el suro que recobreix les parts llenyoses del tronc i l’arrel. També es forma en altres parts de la planta com a conseqüència de les ferides, i actua de teixit cicatritzant. El suro és constituït per diverses capes cel·lulars, però, a diferència de les de l’epidermis, les cèl·lules del suro són mortes, ja que totes les parets cel·lulars són impermeabilitzades per una capa de suberina i, per tant, no són possibles els intercanvis amb l’exterior. Les cèl·lules del suro solen ser plenes d’aire i sovint també contenen tanins. Els tanins són els que donen el color marró o negrós als troncs i a les branques dels arbres. El contingut d’aire de les cèl·lules del suro explica dues propietats típiques d’aquest teixit: la lleugeresa i la capacitat d’actuar com a aïllant tèrmic.
Els teixits parenquimàtics
Els teixits parenquimàtics són els més abundants en les parts toves de la planta. Formen gran part de les fulles, les arrels i les tiges no llenyoses, les llavors i la polpa dels fruits. Són constituïts per cèl·lules vives, fortament vacuolitzades i de forma aproximadament esfèrica o cilíndrica, i sempre presenten espais intercel·lulars.
Els parènquimes tenen diverses funcions importants, entre les quals destaca la fotosíntesi i la reserva. El parènquima especialitzat en la fotosíntesi s’anomena parènquima clorofíl·lic. Les seves cèl·lules tenen una paret prima i nombrosos cloroplasts ben diferenciats.
El parènquima de reserva és format per cèl·lules grosses on s’acumulen substàncies, entre les quals és típic el midó, que la planta utilitzarà en els moments de necessitat. El parènquima en què s’acumula s’anomena amilaci: el citoplasma de les seves cèl·lules conté uns orgànuls anàlegs als cloroplasts (els amiloplasts), on en comptes de clorofil·la hi ha grànuls molt petits de midó. Altres parènquimes de reserva són rics en greixos, proteïnes o sucres. Són molt abundants en òrgans vegetals com les llavors, els tubercles, els bulbs, les arrels i fins i tot les tiges gruixudes; tan sols cal pensar en les moltes substàncies que utilitzem i que provenen de diferens parts de les plantes, com l’oli de llavors, el sucre de canya i de remolatxa, la fècula de patata (midó), etc. El parènquima aqüífer, molt comú en les plantes crasses (els cactus, etc.), també es pot considerar un parènquima de reserva. Les cèl·lules d’aquest parènquima tenen abundància de materials mucilaginosos, els quals posseeixen una capacitat enorme de retenció d’aigua. La presència del parènquima aqüífer, juntament amb el gruix excepcional de la cutícula, proporciona a les plantes crasses una gran capacitat de resistència als climes més àrids.
El parènquima aerífer, particularment desenvolupat en les tiges i les arrels de les plantes aquàtiques o d’aiguamolls, es caracteritza per presentar grans espais intercel·lulars plens d’aire. Les arrels d’aquestes plantes s’enfonsen en el fang, és a dir, en un ambient molt pobre d’oxigen. En aquests casos, l’oxigen arriba a l’arrel des de lluny, des dels òrgans aeris (la tija i les fulles) a través de la xarxa de canals del parènquima aerífer.
Els teixits mecànics o de sosteniment
Els teixits mecànics tenen la funció d’aguantar el pes de la planta i conferir-li resistència als impulsos mecànics (per exemple, les ràfegues de vent). La paret de les seves cèl·lules sempre és molt espessa i sovint lignificada. Es divideixen en colènquimes (les cèl·lules vives de les parets no lignificades) i esclerènquimes (les cèl·lules mortes de les parets lignificades).
Els colènquimes tenen cèl·lules allargades i en les seves parets s’alternen zones gruixudes i zones primes. Aquestes parets tenen poca cel·lulosa i són força deformables. El colènquima és abundant als pecíols de les fulles i a les tiges no llenyoses.
Les cèl·lules dels esclerènquimes són necessàriament mortes perquè la paret gruixuda i lignificada obstaculitza el pas de l’aigua i els soluts. La paret secundària és molt desenvolupada. A diferència dels colènquimes, les parets són elàstiques, però això no vol dir que hagin de cedir.
Les cèl·lules esclerenquimàtiques més importants són les fibres i les esclereides. Les fibres són llarguíssimes (a vegades la llargada es mesura en centímetres), però el seu diàmetre és sempre microscòpic. Tenen forma afuada, punxeguda als extrems, i la paret és tan gruixuda que la llum (cavitat interna d’un òrgan) cel·lular es redueix a un canalet estret. Les fibres solen estar apilades en feixos a la part perifèrica de la tija i estan cimentades per la làmina mitjana. També són molt abundants a les nervadures de les fulles. Les esclereides, menys allargades que les fibres, sovint tenen forma molt irregular amb nombroses protuberàncies i presenten una llum cel·lular més ampla. Moltes vegades es forma una faixa que reforça la perifèria de la tija, on s’alternen grups d’esclereides amb feixos de fibres. Alguns teixits duríssims, com per exemple la capa exterior i llenyosa dels pinyols de les prunes i els albercocs, de les closques de les nous, etc., són fets enterament d’esclereides cimentades.
Els teixits conductors
La funció dels teixits conductors és transportar l’aigua, els ions inorgànics i les molècules orgàniques d’un extrem a l’altre de la planta. Aquest transport funciona en ambdós sentits. L’aigua i els ions inorgànics són absorbits del sòl per l’arrel i transportats cap a les parts aèries (la tija, les fulles, les flors i els fruits). En canvi, les substàncies orgàniques són sintetitzades en les fulles i transportades cap a tots els òrgans no fotosintètics. En un tronc d’arbre els dos corrents de transport viatgen en sentits completament oposats: l’aigua i els ions (saba bruta) pugen cap a les fulles, mentre que les substàncies orgàniques dissoltes en aigua (saba elaborada) baixen cap a les arrels.
Dels dos tipus de transport se n’encarreguen dos teixits diferents: el xilema transporta la saba bruta; el floema, la saba elaborada. Tant el xilema com el floema són teixits complexos, formats per diversos tipus de cèl·lules, i tots dos comprenen elements conductors pròpiament dits, fibres i cèl·lules parenquimàtiques. La velocitat del transport és extraordinàriament diferent en tots dos teixits. La velocitat de pujada es mesura en metres per hora, mentre que la del transport descendent es fa en centímetres per hora, gràcies a la presència de mecanismes eficaços que impedeixen la caiguda per gravetat.
a) El xilema. Els elements conductors del xilema s’anomenen vasos. Són constituïts per cèl·lules allargades, mortes, de les quals només queda la paret. Per tant, recorden tubs buits. La paret és parcialment lignificada, i les zones amb lignina són més gruixudes que la resta de la paret. Aquestes zones engruixides i lignificades de la paret secundària tenen diverses formes: d’anell, d’espiral, d’estructura transversal, etc. En molts casos, aquesta part reforçada de la paret és la més abundant i les zones primes i sense lignificar es redueixen a petites àrees circulars anomenades perforacions. Els porus comuniquen dos elements del xilema (per exemple dos vasos o bé un vas i una cèl·lula parenquimàtica). En realitat, a una perforació d’una determinada zona d’una cèl·lula sempre li correspon una perforació de la cèl·lula veïna, que coincideix exactament amb la primera.
Les perforacions fan de vàlvules, deixen passar l’aigua lliurement però barren el pas de les bombolles d’aire que podrien interrompre l’ascens de l’aigua pels vasos. Els engruiximents dels vasos tenen la funció de donar rigidesa i resistència a les cèl·lules del xilema perquè no s’aixafin, sobretot per a evitar el perill que la cavitat dels vasos es tanqui a conseqüència de les flexions patides per l’òrgan (flexions que es poden verificar per l’acció del vent o els animals, pel pes de la neu, per la pressió del sòl, etc.), amb la qual cosa s’interrompria el flux de la saba.
Hi ha dos tipus de vasos: les traqueides i les tràquees. Les traqueides són cèl·lules llargues i estretes, amb una forma molt semblant a les fibres. L’única diferència és que en aquest cas la llum cel·lular és més ampla, tal com convé a una cèl·lula que ha de transportar material pel seu interior. Cada traqueida comunica amb les de sobre i les de sota a través dels petits porus dels seus extrems. Per tant, una sèrie de traqueides seria comparable a un tub dividit transversalment en diverses seccions. Per contra, les tràquees són comparables a tubs formats d’una sola peça, sense talls transversals. Cada tràquea consta d’una sèrie de cèl·lules superposades les parets transversals de les quals han estat destruïdes totalment o en part durant el desenvolupament. Es per això que les tràquees condueixen l’aigua més ràpidament que les traqueides.
b) El floema. Els elements conductors del floema s’anomenen tubs cribosos. De totes les cèl·lules vegetals són les que divergeixen més de la "cèl·lula estàndard". Entre les seves característiques més notables i curioses destaca el fet que es mantenen vives i funcionals fins i tot sense nucli (que desapareix en una fase molt prematura de la seva vida).
Els tubs cribosos són cèl·lules allargades en el sentit del corrent de transport, generalment amb un calibre menor que el dels vasos del xilema. Les parets transversals de les diverses cèl·lules superposades presenten algunes zones particularment primes, anomenades plaques criboses, perforades per un gran nombre de porus petitíssims a través dels quals passen uns fins cordons citoplasmàtics que comuniquen cada cèl·lula amb les del costat. És a través d’aquests cordons que es transporten les substàncies orgàniques elaborades per les plantes.
Els teixits secretors
Els teixits secretors sintetitzen productes secundaris de diversa mena que queden dins les cèl·lules o que amb més freqüència són rebutjats a l’exterior: olis essencials, resines, gomes, etc.
L’aspecte dels teixits secretors és molt variat. No solen formar grans masses compactes, sinó que es fraccionen en petits grups de cèl·lules, sovint distribuïts per tot el cos de la planta. Són molt comuns els pèls secretors que surten de l’epidermis. La seva cèl·lula terminal engruixida, que recorda el cap d’un llumí, produeix la secreció que s’acumula entre la paret i la cutícula. Els canals secretors, espais intercel·lulars allargats i entapissats per les cèl·lules que hi aboquen la seva secreció, també són molt comuns. En són un exemple els canals resinífers presents en tots els òrgans de les coníferes (el pi, l’avet, etc.).
En el cas dels laticífers, en canvi, les substàncies sintetitzades no s’aboquen a l’exterior. Es tracta de cèl·lules allargades que contenen làtex, un líquid blanquinós, opac i similar a la llet que raja de la planta quan s´hi fa una mínima "ferida". Les plantes que contenen làtex són moltíssimes; entre les més conegudes esmentem la figuera i els seus parents diversos, el pixallits o lletsons i les papaveràcies.
Molts perfums de les plantes són produïts per teixits secretors. Per exemple, el perfum balsàmic, penetrant, de la bosquina mediterrània és degut principalment a la presència d’herbes aromàtiques. Els seus pèls secretors contenen olis essencials que els dies càlids s’evaporen, trenquen la cutícula i s’escampen per l’aire. Gran part de les substàncies aromàtiques emprades a la cuina es troben en els pèls secretors que hi ha a la cara inferior de la fulla de les diverses labiades (la sàlvia, l’orenga, el romaní, l’alfàbrega i la farigola) i als canals secretors de les umbel·líferes (l’api, el fonoll, l’anís, el comí, etc.).