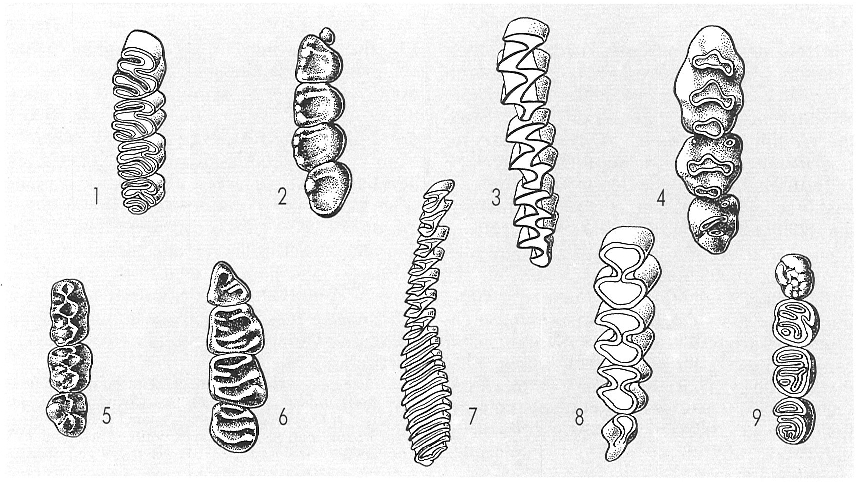
Les corones dentàries dels rosegadors presenten una gran variabilitat, relacionada generalment amb els hàbits alimentaris. El tipus de corona permet agrupar-los en famílies, i les particularitats són emprades en sistemàtica per a diferenciar espècies. L’esquema indica diferents tipus de corona en nou famílies de rosegadors: 1 castòrids, 2 esciúrids, 3 arvicòlids, 4 múrids, 5 cricètids, 6 glírids, 7 hidrocoèrids, 8 octodòntids i 9 histrícids.
Gustavo Hormiga, a partir de fonts diverses.
Els rosegadors constitueixen l’ordre de mamífers més diversificat. Conté unes 1700 espècies vivents, que representen, aproximadament, el 40 % de les espècies de mamífers actuals. No hi ha referències concretes sobre el seu origen evolutiu i, ja des dels fòssils més antics coneguts (Paramys, de finals del Paleocè o principis de l’Eocè, fa aproximadament uns cinquanta milions d’anys), es presenten caràcters comuns a tots els rosegadors: un parell d’incisives amb creixement continu, presència d’un diastema, absència de canines i petites dimensions del cos. Això apunta vers un origen monofilètic del grup, que hauria sorgit de mamífers placentaris primitius mitjançant un procés desconegut fins ara. No sabem ni on ni com tingué lloc aquest procés. Des de l’Eocè, els rosegadors presentaren un procés evolutiu ràpid i simultani en moltes línies, la majoria de les quals han persistit fins avui. Aquest procés és simultani a la gran expansió que efectuaren fins arribar a ocupar tots els continents, excepció feta d’Austràlia (on penetraren molt més tard, a finals del Terciari, fa aproximadament dos o tres milions d’anys).

Els rosegadors constitueixen el grup més diversificat dels mamífers i han colonitzat un gran nombre de medis (subterrani, arbori, desèrtic, estèpic, etc.) adoptant morfologies adequades a cada cas. A la figura s’exposa un exemple de les formes més comunes: 1 rata, 2 jerbu, 3 llebre de Patagònia, 4 espàlax, 5 paca, 6 esquirol, 7 porc espí, 8 capibara.
Gustavo Hormiga, a partir de fonts diverses.
La seva radiació evolutiva es caracteritza perquè presenta, en lloc de l’aspecte arborescent típic de molts grups zoològics, una representació a partir de línies paral·leles, les relacions de les quals a l’origen són difícils d’establir. Aquest gran nombre de paral·lelismes dificulta molt la interpretació dels caràcters a l’hora d’intentar establir relacions filogenètiques dins de l’ordre. Les característiques estructurals dels rosegadors primitius, que van assegurar el seu gran èxit evolutiu, van limitar, també, el desenvolupament d’aquest grup. La gran majoria han quedat com a formes petites i herbívores, sense modificacions particulars de les extremitats. Aquesta restricció del seu procés evolutiu, juntament amb el desenvolupament d’una gran riquesa de formes, ha produït que l’evolució paral·lela s’hagi donat més entre els rosegadors que entre qualsevol altre grup de mamífers. Moltes formes no relacionades entre elles han desenvolupat els mateixos trets i han esdevingut similars per paral·lelisme. Les tendències evolutives, que apareixen independentment en els diferents grups de rosegadors, inclouen: el desenvolupament de diferents tipus de masseters que han donat lloc als tres tipus principals, l’esciuromorf, l’histricomorf i el miomorf (emprats com a criteris taxonòmics); la reducció de les premolars: mai no n’hi ha més de dues en una sèrie dentària i, sovint, desapareixen del tot (normalment, hi ha tres dents molariformes); la hipsodòncia, o creixement de les dents en longitud, que condueix, sovint, a la desaparició de les arrels i a les dents de creixement continu; la complicació de l’estructura dentària per l’aparició de cúspides addicionals que formen carenes d’esmalt, o, alternativament, la simplificació de l’estructura dentària, per causa del creixement continu, fins que les dents queden convertides en simples pilars.
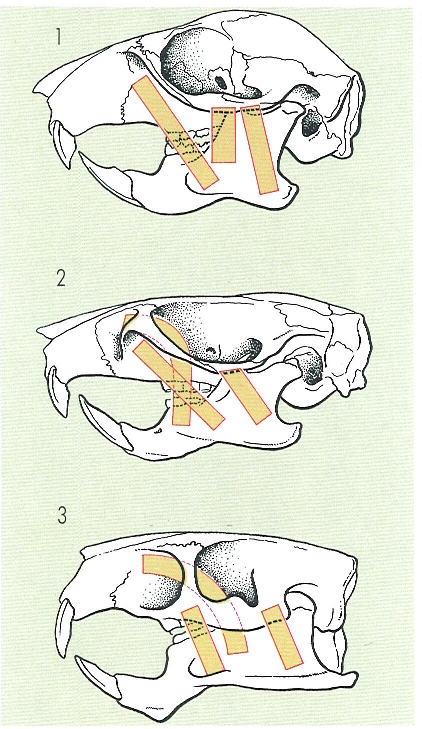
La disposició dels masseters als rosegadors determina, malgrat la dificultat d’establir-hi línies evolutives, les tres possibles vies que, a grans trets, s’accepten dins d’aquest grup: 1 disposició esciuromorfa, 2 disposició miomorfa i 3 disposició histricomorfa.
Gustavo Hormiga, a partir de fonts diverses.
Els trets més diferencials de l’ordre es troben a la seva dentició. Presenten un sol parell d’incisives a cada hemimandíbula, cosa que els diferencia, entre altres coses, dels lagomorfs, que en tenen dos. Aquestes dents tenen esmalt, només, a la superfície anterior i la resta és formada per dentina que, en desgastar-se, genera la característica cara bisellada del seu extrem. Són de creixement continu i les arrels són obertes. Entre aquestes dents i les molariformes (premolars, en cas que hi hagi molars), hi ha un ampli espai lliure, que és el diastema. No presenten canines i les premolars hi són en nombre reduït (una o dues) o arriben a desaparèixer. Les molars poden tenir creixement continu o no, segons el grup; presenten gran diversitat morfològica i la corona pot adoptar formes diverses que, a diferents nivells, són emprades en sistemàtica.
Els rosegadors presenten una àmplia gamma de formes i adaptacions. La seva grandària oscil·la entre els 5-7 cm del ratolí espiguer d’Europa i els 125 cm (amb un pes de 50 kg) de la capibara d’Amèrica del Sud (Hydrochoerus hydrochoeris). La morfologia més general és la que correspon a formes de dimensions petites (8-10 cm del ratolí domèstic) o mitjanes (25-30 cm de les rates) amb el cos allargat, musell relativament prominent, potes curtes i cua llarga, com les rates i ratolins, o curta com els talpons. Sobre aquesta base morfològica més freqüent, hi ha una àmplia gamma de variacions adaptatives a condicions particulars.
La majoria d’espècies són terrestres, però també n’hi ha que porten una vida subterrània, arborícola o aquàtica i presenten adaptacions particulars en cada cas. Les subterrànies (Spalax) tendeixen a tenir el cos cilíndric, les potes curtes i robustes, i la cua, les orelles i els ulls rudimentaris. La vida aquàtica comporta la presència de membranes interdigitals, més o menys desenvolupades, i l’aplatament (dorsiventral en el castor i lateral en la rata mesquera) de la cua per a facilitar la natació. Els de costums arborícoles presenten la cua ben desenvolupada, sovint peluda (esquirol), que ajuda a mantenir l’equilibri durant el salt; de vegades, es desenvolupa una membrana entre les extremitats que permet planar a l’animal (esquirol volador). Els rosegadors de zones obertes, deserts i estepes s’han adaptat al desplaçament ràpid mitjançant grans salts. Les extremitats posteriors són ben desenvolupades i les anteriors són curtes, els dits externs dels peus es redueixen i la cua s’allarga (el mateix que succeeix en els cangurs); tots aquests tipus d’adaptacions es poden donar en diferents línies evolutives de rosegadors, de tal manera que es produeixen, independentment i per paral·lelisme, convergències de forma.
L’existència d’autotomia caudal en els rosegadors és un fenomen rar entre mamífers. Es dóna, per exemple, en el ratolí lleonat (Apodemus flavicollis) i en el liró. Consisteix en el fet que la coberta de la part terminal de la cua es desprèn amb facilitat, deixant les vèrtebres al descobert. L’hemorràgia de la zona despullada és molt dèbil i, al cap d’un temps, s’asseca i cau. No hi ha regeneració.
El nombre de mamelles varia; sovint se’n troben diversos parells (fins onze), des del pit fins a l’àrea inguinal. Les mans i els peus són generalment pentadàctils; els dits externs poden reduir-se, especialment els de les formes saltadores, i algunes espècies tenen el primer dit oposable com a adaptació per a agafar-se a les branques.
El crani és normalment baix, allargat i aplatat dorsiventralment. La cavitat encefàlica és petita. L’òrbita ocular sempre és oberta i presenten una arcada zigomàtica ben desenvolupada. El forat suborbital, en el maxil·lar, és gran i una part del masseter pot passar-hi a través; l’os timpànic sempre constitueix una bul·la timpànica, que, ocasionalment (especialment en els rosegadors del desert), és molt gran. La mandíbula inferior és robusta i les dues hemimandíbules mai no es fonen, sinó que són unides per un lligament que els permet un cert grau de mobilitat, la qual es manifesta durant la prensió i la masticació de l’aliment. La mandíbula és capaç de realitzar moviments anteroposteriors i laterals; d’aquesta manera, l’aliment es tritura mitjançant moviments circulars. Amb l’excepció del crani, l’esquelet dels rosegadors és bastant uniforme.
Els hemisferis cerebrals són, generalment, llisos i només en espècies grans es perceben dèbils circumvolucions. L’olfacte és el sentit bàsic dels rosegadors, fet que s’evidencia pel gran desenvolupament que presenten els lòbuls olfactius. L’oïda té un paper important en les formes desèrtiques, on les densitats de població són baixes i els animals es troben molt disseminats, ja que facilita les possibilitats de relació.
Els rosegadors són generalment herbívors, si bé hi ha espècies que també inclouen animals a la seva dieta. Moltes espècies presenten bosses a les galtes que empren per emmagatzemar i transportar l’aliment (com els hàmsters).
En el cas dels mascles, els testicles creixen notablement de dimensions durant el període reproductor. Només durant aquest període es desplacen des de la cavitat abdominal fins a sota la pell de l’abdomen o l’escrot per tal que l’espermatogènesi sigui possible. En les femelles, l’úter és dividit en diferents graus, encara que generalment és de tipus bicorne. L’ovulació és espontània, però en algunes espècies té lloc només després de la còpula. La majoria de rosegadors són polièstrics. En climes temperats i freds s’estableix una època de descans reproductor, que sol coincidir amb l’època més freda. En els animals antròpics, com per exemple el ratolí domèstic o la rata comuna, les poblacions que viuen a l’exterior dels habitacles humans segueixen el cicle reproductor en funció de les condicions ambientals; en canvi, a les poblacions que viuen a ciutats, granges, etc., com que ni l’aliment ni l’ambient no són factors limitants, el cicle reproductor es perllonga durant tot l’any. Més que els factors ambientals (com, per exemple, la temperatura), sembla que el factor limitant més important és la disponibilitat d’aliment.
El període de gestació és relativament llarg; al voltant de 20 dies, el del ratolí domèstic, i 150 dies el del coipú. El nombre de cries per gestació és, també, relativament gran; una rata comuna o un hàmster poden arribar a tenir divuit cries per part, i els petits de diverses espècies neixen amb diferents graus de desenvolupament. En les espècies petites, particularment en els talpons, els joves arriben a la maduresa sexual molt aviat. La vida mitjana d’un rosegador petit és curta; els talpons i ratolins rarament ultrapassen l’any de vida.
En la seva majoria són nocturns. Les espècies que es nodreixen de substàncies de poder nutritiu baix (cas dels talpons, que mengen herba) han d’omplir l’estómac diverses vegades durant el dia i la nit i, per això, els períodes de descans són curts i se’ls pot veure durant el dia; però, malgrat tot, són més actius durant les hores nocturnes. Les espècies de regions fredes (al N i a l’alta muntanya) tendeixen a ésser actives durant el dia, en canvi, les desèrtiques surten de nit. A les regions on hi ha una estació freda marcada, hi ha espècies que tenen hivernació (cas de la marmota, lirons, etc.) i, en canvi, a les regions amb una estació marcadament seca, hi ha espècies que tenen estivació (esquirols de l’estepa, per exemple). Moltes espècies excaven galeries, on es refugien dels seus enemics, s’amaguen durant el repòs, hivernen o crien. Altres construeixen nius, i els castors, a més d’excavar caus, aixequen dics per crear estanys al voltant dels seus caus.
L’ordre dels rosegadors és cosmopolita. Les formes sinantròpiques, com els ratolins i les rates, són presents a totes les zones habitades per l’home. La capacitat de colonització i d’adaptació dels ratolins és molt elevada.
Segons Kowalski (1981), durant el Terciari inferior els rosegadors eren notablement diversificats geogràficament, de manera que molts grups de rosegadors fòssils eren ubicats en regions concretes. Posteriorment, les connexions intercontinentals facilitaren certes barreges. A l’Oligocè (fa uns 35 milions d’anys), els caviomorfs començaren a desenvolupar-se a Amèrica del Sud. Aquest grup romangué molt de temps restringit a aquest continent, on arribà a diversificar-se molt. Només a finals del Terciari, després de formar-se la unió continental entre Amèrica del Nord i Amèrica del Sud, els cricètids i esciúrids d’Amèrica del Nord es desplaçaren vers el S, i els caviomorfs ho feren vers el N. Els altres grups de rosegadors s’originaren en diferents continents. Els grups adaptats a un clima fred s’estengueren entre Euràsia i Amèrica del Nord (castòrids, esciúrids, cricètids); els d’origen més meridional estigueren més restringits en la seva expansió. Els geòmids i els heteròmids es troben restringits a Amèrica. Els múrids, glírids, dipòdids, espalàcids, rizòmids i histrícids són presents només a l’antic continent. Els trionomioïdeus, anomalúrids, pedètids i batièrgids semblen ésser grups típicament africans.
Tenint en compte el paral·lelisme evolutiu i la convergència que ja hem esmentat, i també el coneixement molt fragmentari dels fòssils del període en què els rosegadors es diferenciaren, l’ordenació taxonòmica dels representants de l’ordre és molt problemàtica. L’any 1855, Brand ordenà els rosegadors en tres subordres: els esciuromorfs, els histricomorfs i els miomorfs, a partir de l’estructura dels masseters. Aquesta ordenació encara s’empra avui, especialment en treballs sobre sistemàtica de rosegadors actuals; no obstant això, no hi ha dubte que no reflecteix les relacions naturals entre els grups (un grup inclou els porcs espins de l’antic continent i els caviomorfs d’Amèrica del Sud, que s’han desenvolupat, òbviament, per separat). Intents posteriors de classificació tampoc no han solucionat el problema, si bé la tendència actual és mantenir el subordre dels esciuromorfs, separar els glírids dels miomorfs, establint un nou subordre, els gliriomorfs, i escindir el subordre dels histricomorfs en dos, mantenint el terme "histricomorfs" per als representants de l’antic continent i agrupant tots els abans denominats histricomorfs del nou continent dins del subordre dels caviomorfs. Thenius, en un estudi en què fa una revisió profunda basada en restes fòssils, suggereix i justifica l’increment de subordres tot separant dels esciuromorfs els castorimorfs i els anomaluromorfs, a més dels gliriomorfs. En canvi, aquest autor considera que no hi ha prou justificació per a escindir els caviomorfs dels histricomorfs. A la present obra, hem adoptat l’estructura de subordres de Thenius tot mantenint separats els caviomorfs. L’agrupació en superfamílies l’hem feta d’acord amb Kowalski i les famílies considerades, en funció de la revisió més recent de Honacki, Kinman i Koeppl.
A tot el món, hi ha 1700 espècies vivents, distribuïdes en 390 gèneres i 33 famílies. Als Països Catalans, n’hi ha 18 espècies (un 1 % del total mundial), pertanyents a quatre famílies, que tractarem a continuació.
A continuació, la taula mostra la distribució de l’ordre dels rosegadors a la península Ibèrica i als Països Catalans. PPCC Països Catalans, CN Catalunya Nord, A Andorra, P Principats, PV País Valencià, MA Mallorca, ME Menorca, El Eivissa, FO Formentera, CA Cabrera, PI Península Ibèrica, F França; + espècie autòctona, i introduïda recentment,? de presència no comprovada, * escapada de granges de cria. (Dades elaborades per l’autor).
Distribució de l’ordre dels rosegadors a la península Ibèrica i als Països Catalans
| ESPÈCIE | PPCC | CN | A | P | PV | MA | ME | El | FO | CA | PI | F | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ESCIÚRIDS | |||||||||||||
| Esquirol (Sciurus vulgaris) | + | + | + | + | + | – | – | – | – | – | + | + | |
| Marmota (Marmota marmota) | i | – | – | – | – | – | – | – | – | – | i | + | |
| CASTÒRIDS | |||||||||||||
| Castor (Castor fíber) | – | – | – | – | – | – | – | – | – | – | – | + | |
| GLÍRIDS | |||||||||||||
| Rata cellarda (Eliomys quercinus) | + | + | + | + | + | + | + | – | + | – | + | + | |
| Liró gris (Glis glis) | + | + | + | + | – | – | – | – | – | – | + | + | |
| Muscardinus avellanarias | – | – | – | – | – | – | – | – | – | – | – | + | |
| MÚRIDS | |||||||||||||
| Ratolí espiguer (Micromys minutus) | + | + | – | – | – | – | – | – | – | – | + | + | |
| Ratolí de bosc (Apodemus sylvaticus) | + | + | + | + | + | + | + | + | + | – | + | + | |
| Ratolí gorjagroc (A. flavicollis) | ? | ? | ? | – | – | – | – | – | – | – | + | + | |
| Rata negra (Rattus rattus) | + | + | + | + | + | + | + | + | + | + | + | + | |
| Rata comuna (R. norvegicus) | + | + | + | + | + | + | + | + | + | + | + | + | |
| Ratolí domèstic (Mus musculus) | + | + | + | + | + | + | + | + | + | + | + | + | |
| Ratolí mediterrani (M. spretus) | + | + | – | + | + | + | + | + | – | – | + | + | |
| ARVICÒLIDS | |||||||||||||
| Talpó roig (Clethrionomys glareolus) | + | + | + | + | – | – | – | – | – | – | + | + | |
| Rata talpera (Arvicola terrestris) | + | + | – | + | – | – | – | – | – | – | + | + | |
| Rata d’aigua (A. sapidus) | + | + | + | + | + | – | – | – | – | – | + | + | |
| Rata mesquera (Ondatra zibethicus) | – | – | – | – | – | – | – | – | – | – | – | + | |
| Talpó comú (Microtus duodecimcostatus) | + | + | – | + | + | – | – | – | – | – | + | + | |
| Talpó pirinenc (M. Pyrenaicus) | + | + | + | + | – | – | – | – | – | – | + | + | |
| M. lusitanicus | – | – | – | – | – | – | – | – | – | – | + | + | |
| M. subterraneus | – | – | – | – | – | – | – | – | – | – | – | + | |
| M. multiplex | – | – | – | – | – | – | – | – | – | – | – | + | |
| Talpó de tartera (M. nivalis) | + | + | + | + | – | – | – | – | – | – | + | + | |
| Talpó dels prats (M. arvalis) | + | + | + | + | – | – | – | – | – | – | + | + | |
| Talpó muntanyenc (M. agrestis) | + | + | + | + | – | – | – | – | – | – | + | + | |
| Talpó de Cabrera (M. cabrerae) | – | – | – | – | – | – | – | – | – | – | + | + | |
| CAPRÒMIDS | |||||||||||||
| Coipú (Myocastor coipus) | * | – | – | * | – | – | – | – | – | – | * | i | |
| TOTAL | 19 | 18 | 13 | 16 | 9 | 6 | 6 | 5 | 5 | 3 | 21 | 27 |
La classificació dels rosegadors es presenta a la taula següent: relació dels subordres, les superfamílies i les famílies dels rosegadors actuals. Amb un interrogant (?) s’indiquen les famílies la relació de les quals amb alguna superfamília encara no s’ha pogut establir. Amb un asterisc (*) s’indiquen les que tenen representació als Països Catalans; assenyalem a continuació el nombre d’espècies que hi són presents: esciúrids, 2; glírids, 2; arvicòlids, 8; i múrids, 6. (Dades elaborades per l'autor).
| O. ROSEGADORS | ||
|---|---|---|
| SO. protrogomorfs (Protrogomorpha) | ||
| SpF. aplodontoïdeus (Aplodontoidea) | ||
| F. aplodòntids (Aplodontidae) | ||
| SO. esciuromorfs (Sciuromorpha) | ||
| SpF. esciuroïdeus (Sciuroidea) | ||
| F. esciúrids (*) | ||
| SpF. geomioïdeus (Geomyoidea) | ||
| F. geòmids (Geomydae) | ||
| F. heteròmids (Heteromyidae) | ||
| SpF. diplodoïdeus (Diplodoidea) | ||
| F. dipòdids (Dipodidae) | ||
| F. zapòdids (Zapodidae) | ||
| SO. castorimorfs (Castorimorpha) | ||
| SpF. castoroïdeus (Castoroidea) | ||
| F. castòrids (Castorídae) | ||
| SO. anomaluromorfs (Anomaluromorpha) | ||
| SpF.? | ||
| F. anomalúrids (Anomaluridae) | ||
| F. pedètids (Pedetidae) | ||
| SO. gliriomorfs (Gliriomorpha) | ||
| SpF. gliroïdeus (Gliroidea) | ||
| F. glírids (*) | ||
| F. selevínids (Seleviniidae) | ||
| SO. miomorfs (Myomorpha) | ||
| SpF. muroïdeus (Muroidea) | ||
| F. cricètids (Crícetidae) | ||
| F. arvicòlids (Arvicolidae) (*) | ||
| F. múrids (Muridae) (*) | ||
| SpF. espalacoïdeus (Spalacoidea) | ||
| F. espalàcids (Spalacidae) | ||
| F. rizòmids (Rhizomyidae) | ||
| SO. histricomorfs (Hystricomorpha) | ||
| SpF. histricoïdeus (Hystrícoidea) | ||
| F. histrícids (Hystricidae) | ||
| SpF. trionomioïdeus (Thryonomyoidea) | ||
| F. trionòmids (Thryonomyidae) | ||
| F. petromúrids (Petromuridae) | ||
| SpF. batiergoïdeus (Bathyergoidea) | ||
| F. batièrgids (Bathyergidae) | ||
| SpF.? | ||
| F. ctenodactílids (Ctenodactylidae) | ||
| SO. caviomorfs (Caviomorpha) | ||
| SpF. octodontoïdeus (Octodontoidea) | ||
| F. octodòntids (Octodontidae) | ||
| F. ctenàmids (Ctenamyidae) | ||
| F. equímids (Echimyidae) | ||
| F. capròmids (Capromyidae) | ||
| F. abrocòmids (Abrocomyidae) | ||
| SpF. xinxil·loïdeus (Chinchilloidea) | ||
| F. xinxíl·lids (Chinchillidae) | ||
| SpF. cavioïdeus (Cavioidea) | ||
| F. càvids (Caviidae) | ||
| F. hidroquèrids (Hydrochoeridae) | ||
| F. dinòmids (Dinomyidae) | ||
| F. dasipròctids (Dasiproctidae) | ||
| F. agoútids (Agoutidae) | ||
| SpF. eretizontoïdeus (Erethizontoidea) | ||
| F. eretizòntids (Erethizontidae) | ||