Les estratègies de supervivència en l’aridesa
L’aridificació dels climes de les regions tropicals i subtropicals que avui ocupen deserts i subdeserts càlids sembla que començà durant el Miocè. En aquelles dates el continent australià ja estava aïllat, cosa que explicaria la singularitat de la seva fauna amb independència de les condicions bioclimàtiques presents. El mateix passava amb els dos subcontinents americans que, a més, estaven separats entre ells i hi romangueren fins que, al Pliocè, es formà l’istme de Panamà i es féu possible un cert intercanvi de faunes. També fou durant el Miocèque la placa continental afro-aràbiga col·lisionà ambla centrasiàtica fent desaparèixer l’antiga mar de Tetis i unificant les àrees subtropicals del que avui és el Vell Món. Això explica la relativa unitat de les faunes desertícoles d’aquests darrers, la dels deserts i els subdeserts nord i sud-americans, bé que amb peculiaritats indubtables d’uns i altres, i la total diferenciació entre la fauna dels deserts australians i la de tots els altres. El resultat de tots els trasbalsos i de la progressiva adaptació a les condicions de vida dels deserts dels animals amb menys mobilitat són les respectives faunes presents avui a les diferents àrees de deserts i subdeserts.
Els animals del desert s’enfronten a un problema ambiental comú, l’escassetat d’aigua, agreujat per les altes temperatures, que n’incrementen l’evaporació. Els animals perden aigua principalment a través de la transpiració, la respiració i l’excreció. Per tal de subsistir amb una ingesta d’aigua mínima, n’han de reduir les pèrdues, i l’han de conservar costi el que costi. Això s’aconsegueix mitjançant l’ús, generalment coordinat, de nombrosos mecanismes morfològics, fisiològics i etològics.
Les diferents formes de vida latent i de dormència
Els animals petits, com els insectes, sovint poden defugir la secada i la calor del desert completant els seus cicles vitals durant l’estació de les pluges, abans del començament de les condicions extremes: aquestes se suporten en un estat fisiològic de mort aparent conegut com a diapausa. Aleshores el metabolisme es redueix a un nivell baix, de tal manera que es redueixen al mínim les pèrdues en la respiració. Els ous d’alguns crustacis, com les tortuguetes (Triops), i també els de molts insectes, sobreviuen a la calor i la sequedat de l’estiu del desert en diapausa. Aquests ous poden tolerar tant les temperatures extremament altes com les baixes i són molt resistents a la pèrdua de vapor d’aigua per transpiració.
Per altra banda, molts animals del desert més grans eviten les condicions adverses de la secada i la calor restant en un estat quiescent. Tanmateix, en el seu cas, no es produeix la diapausa, ja que en l’estivació no hi ha hormones involucrades en el control del procés i els animals es reactiven així que les condicions milloren. Durant l’estivació, l’obertura de la closca dels cargols del desert, per exemple, és closa per un gruixut tel de secreció mucosa calcificada anomenat epifragma, que redueix la pèrdua d’aigua per evaporació fins a tal punt que se sap que alguns d’aquests animals han romàs en aquest estat durant més de cinc anys i s’han recuperat tan bon punt s’han mullat. El seu baix ritme metabòlic assegura la supervivència durant llargs períodes de letargia.
Altres animals, com els amfibis, estiven en capolls de fang sec o pell morta profundament enterrats i relativament impermeables a l’aigua. Els animals queden en un estat de mort aparent, durant el qual el ritme metabòlic es redueix moltíssim, i depenen de les reserves de greix que tenen a la cavitat abdominal. Els gripaus d’esperons americans (Scaphiopus), per exemple, romanen inactius durant la major part de l’any, i estiven fins a 1 m per sota de la superfície del sòl. Cada estiu surten la primera nit de pluges fortes (a Arizona, per exemple, generalment al principi del juliol) i ponen els ous als tolls acabats de formar. El període subsegüent de condicions favorables persisteix durant poc mésde vuit setmanes, època durant la qual els caps-gros-sos s’han de desenvolupar ràpidament i els gripaus adults han de menjar prou per a poder reposar les seves reserves de greix abans del següent llarg període d’estivació. En condicions naturals, són estimulats a sortir del seu cau pel so produït pels trons i les gruixudes gotes de pluja dels aiguats. Malauradament, també responen a les vibracions produïdes per les motos i altres vehicles mecànics que travessen el desert, cas en el qual, si són induïts a sortir dels seus capolls abans del començament de l’estació de les pluges, quan no hi ha menjar ni aigua disponible, experimenten un gran trasbals, no tenen prou reserves per tornar a refugiar-se sota terra i es moren. Poblacions senceres de gripaus d’esperons americans poden ser exterminades accidentalment d’aquesta manera.
El control tèrmic
Evitar i defugir la calor i la secada s’aconsegueix mitjançant característiques morfològiques i fisiològiques que redueixen l’augment de la calor i la pèrdua d’aigua, i mitjançant estratègies de comportament, com ara amagar-se en caus i recers durant les hores més caloroses del dia. La majoria de llangardaixos del desert, per exemple, són actius durant el dia, però mantenen una temperatura corporal extraordinàriament constant movent-se del sol a l’ombra.
Tots els aspectes de l’ecofisiologia estan interrelacionats, però en els calorosos deserts cap no ho està més que les relacions tèrmiques i hídriques. Com que el poder dessecant de l’aire s’incrementa tant a temperatures altes, la calor i la secada són de bon tros els dos elements més importants del clima. Tant els artròpodes com els rèptils estan dotats d’unes superfícies externes relativament impermeables. Tot i que la pèrdua cutània d’aigua en els rèptils del desert és lleugerament més alta del que s’havia cregut, és evident que és considerablement més baixa que no pas en espècies d’entorns més humits. En la major part dels casos se sap poc sobre els factors que hi intervenen, però sembla que la presència d’escates no és significativa. Es va descobrir que els índexs de pèrdua d’aigua d’una mutació sense escates de la serp nord-americana Pituophis melanoleucus eren comparables amb els d’un individu normal, malgrat el fet que a la serp sense escates fins i tot li mancava la capa externa de pell superficial, i que tenia una capa de ceratina molt més prima del que és normal.
Una capa de sorra humida acumulada sobre els narius pot reduir la pèrdua respiratòria d’aigua humitejant l’aire inspirat i atrapant una part del vapor d’aigua de l’aire expirat. Aquesta és l’estratègia que fan servir molts llangardaixos excavadors, com l’Aporosaura anchietae, que eviten l’entrada de grans de sorra a l’interior dels narius mitjançant unes vàlvules nasals. Les tortugues retenen la respiració quan perden aigua, mentre que els jerbus, les rates cangur i altres petits rosegadors del desert tenen uns conductes nasals estrets amb una gran àrea superficial que es refresca amb l’evaporació quan l’aire és inspirat: aleshores l’aigua es torna a condensar durant l’expiració. La gariba (Echis carinatus), la cerasta o escurçó banyut (Cerastes cerastes) i l’escurçó saharià (C. vipera) estridulen refregant les puntes de les seves escates laterals i eviten així la pèrdua d’humitat respiratòria que els ocasionaria xiular.
Per als vertebrats més grans, sobretot ocells i mamífers, la pèrdua d’aigua cutània no és tan important com en les sargantanes i serps petites. Això és perquè quan el volum d’un cos es cubica, la seva àrea superficial només s’eleva al quadrat. Com més gran és un animal, més petita és, relativament, la seva àrea superficial. Aquesta simple relació explica les llargues cames de les ovelles i les cabres del desert comparades amb les de les varietats de climes més frescos. Així i tot, només els mamífers del desert comparativament grans tenen una proporció entre superfície i volum prou petita que els permet refrescar els seus cossos mitjançant l’evaporació de la suor. Com que tenen una reserva d’aigua més gran per unitat d’àrea superficial, aquests animals grans, per exemple el camell comú o dromedari (Camelus dromedarius), l’ase (Equus asinus), l’addax (Addax nasomaculatus), els òrix (Oryx) o la gasela comuna (Gazella dorcas), poden resistir més temps que els mamífers més petits, com els jerbus o les rates cangur, un cert índex de pèrdua d’aigua sense que s’arribi a nivells letals.
La pèrdua de calor a través de la suor en els vertebrats més grans no es produeix fins que la temperatura corporal no s’eleva molt per damunt del que és normal. D’aquesta manera, la calor s’emmagatzema durant el dia i es perd a la nit quan la temperatura ambiental baixa; així s’evita una pèrdua d’aigua excessiva. Els camells, per exemple, no comencen a suar fins que la seva temperatura corporal no s’ha elevat fins a 40,7°C. A més a més, quan la temperatura del camell augmenta, la diferència entre aquesta i la de l’aire que l’envolta es redueix, de tal manera que es necessita menys suor per a evitar un augment més gran de la temperatura corporal. D’altra banda, l’aspre pèl del seu llom actua com una barrera contra la radiació solar i alenteix la conducció de la calor ambiental. Les temperatures corporals elevades també es poden observar en l’addax, l’òrix, la gasela comuna o l’estruç. Tot i que les temperatures corporals de la gasela i l’antílop poden arribar als 46°C, el subministrament de sang al cervell es refreda per l’intercanvi de calor al sinus carotidi, una xarxa de petits vasos sanguinis del sinus cavernós. Aquest sí que és ple de sang venosa que prové dels conductes nasals, on ha estat refredada per l’evaporació de les humides membranes mucoses.
Tot i que els ocells no suen, perden calor i humitat alenant i durant el vol, quan l’aire és impulsat cap als sacs aeris a través dels pulmons. Com a conseqüència, una quantitat relativament gran d’aigua es perd a través de l’evaporació i els ocells petits que són, en general, menys eficients que els mamífers petits en els medis àrids, han de beure per tal de sobreviure, però això els ho facilita la seva considerable mobilitat. Els rapinyaires més grans poden tenir-ne prou amb els fluids corporals de la seva presa.
L’optimització fisiològica de la ingestió d’aigua
Una adaptació fisiològica comuna entre els herbívors i els carnívors del desert és la capacitat de viure de l’aigua obtinguda a través del menjar. Els insectes tendeixen a conservar els alts continguts d’aigua de les plantes i els seus cossos poden contenir una quantitat d’aigua superior a la que es troba a la seva dieta. Les gaseles i els antílops pasturen a la nit menjant fulles que transpiren activament i que contenen grans quantitats d’aigua. Aixoplugades als seus caus relativament frescos i humits durant el dia, les rates cangur i els jerbus poden mantenir l’equilibri d’aigua amb una dieta de llavors seques, sense beure mai, ja que si haguessin de respirar l’aire sec de fora dels seus caus durant el dia, l’índex d’evaporació dels seus pulmons excediria l’índex de formació d’aigua metabòlica. Altres mamífers del desert també poden obtenir suficient humitat per a les seves necessitats a través de l’aigua present en els seus aliments, com la rata de sorra (Psammomys obesus), que s’alimenta de plantes suculentes, les rates tragineres americanes o “pack-rats” (Neotoma), els esquirols de terra (Spermophilus [=Citellus]), que s’alimenten dels sucosos fruits de la “cholla” o nopal (Opuntia) o els ratolins llagosters (Onychomys), amb una dieta insectívora que cobreix totes les seves necessitats d’aigua. La “mulgara” (Dasycercus cristicauda) també s’alimenta sobretot d’insectes i excreta només un petit volum d’orina molt concentrada. Els grans mamífers, a més d’altres sistemes ja esmentats, també intenten reduir les pèrdues d’aigua mitjançant la producció d’orina molt concentrada, d’excrements deshidratats i la ingesta de les plantes comestibles amb les fulles més humides.
Els productes d’excreció d’insectes i aràcnids, àcid úric i guanina respectivament, són summament insolubles. Com a conseqüència, els residus nitrogenats es poden eliminar del cos en un estat sec i sense perdre aigua en el procés. Els residus urinaris dels rèptils i els ocells s’eliminen com una massa polposa o semisòlida que conté una elevada proporció d’àcid úric i molt poca aigua. L’excreció d’àcid úric és molt avantatjosa ja que es pot formar orina que té una salinitat més elevada que la del plasma sanguini i va associada amb la posta d’ous en medi aeri tancats en unes membranes relativament impermeables. Si un ocell o un rèptil embrionaris produïssin amoníac, tal com fan els peixos, s’enverinarien. La urea, pròpia dels mamífers, com que és soluble, podria arribar a alterar l’equilibri osmòtic intern de l’ou en assolir concentracions massa elevades i causar així la mort de l’embrió. L’àcid úric, en canvi, es pot deixar enrere, dins de la closca, quan el jove insecte, rèptil o ocell la trenca i surt de l’ou. Desenvolupada pels embrions, l’excreció d’àcid úric ha estat mantinguda pels animals adults i és especialment útil en regions on l’aigua és escassa. Gràcies a aquestes adaptacions, insectes, aranyes i escorpins del desert són extremament econòmics pel que fa a l’ús de l’aigua.
Les proteccions cuticulars i anatòmiques dels artròpodes
Amb poques excepcions, els artròpodes del desert que no tenen cicles vitals curts amb diapausa sobreviuen a la calor i la secada diürnes vivint sota roques o pedres, o en profunds caus excavats a la sorra. A causa de la seva mida reduïda, la seva proporció entre superfície i volum és extremament alta, i per tal de poder sobreviure al desert (de fet, a qualsevol entorn terrestre que no sigui molt humit) han de posseir necessàriament una coberta que sigui ben resistent al pas de l’aigua. Fins i tot les pasteretes, que són crustacis isòpodes —la majoria viuen en medis força més humits—, han desenvolupat als deserts cutícules sorprenentment resistents a les pèrdues d’aigua. Gràcies a les seves adaptacions etològiques, per exemple, Hemilepistus reaumuri, una d’aquestes pasteretes o porquets de Sant Antoni, és un dels herbívors i detritívors més resistents de la macrofauna de moltes zones àrides de l’Àfrica septentrional i del Pròxim Orient.
Els aràcnids i els insectes posseeixen unes cutícules fins i tot més resistents a la pèrdua d’aigua per transpiració que les pasteretes del desert. La capa externa del tegument, l’epicutícula, conté una capa monomolecular de lípid a la qual el tegument deu pràcticament tota la resistència a la pèrdua d’aigua. La temperatura a la qual les molècules de la capa de cera permeten el pas del vapor d’aigua és molt més alta en els insectes del desert i els aràcnids que no pas en espècies comparables d’entorns temperats.
Un artròpode enterament cobert amb una capa de cera epicuticular gairebé impermeable gaudiria de l’estimable avantatge de perdre molt poca aigua en la transpiració, però en canvi no podria respirar: la molècula d’oxigen és, tanmateix, més gran que les molècules d’aigua. Així, han hagut d’evolucionar mecanismes respiratoris que permetessin l’intercanvi gasós sense pèrdues d’aigua excessives. En el cas de miriàpodes i insectes, un sistema d’espiracles, tràquees i traquèoles transporta l’oxigen directament als teixits on tenen lloc els processos metabòlics. En la major part dels aràcnids, per la seva banda, els principals òrgans respiratoris són sacs pulmonars que comuniquen amb l’atmosfera externa a través d’uns petits orificis i contenen nombroses lamel·les respiratòries. Els escorpins només tenen sacs pulmonars, mentre que les aranyes posseeixen tant sacs pulmonars com tràquees, poc significatives pel que fa a la respiració. Les tràquees han evolucionat de manera secundària en els solífugs, i la seva presència està relacionada amb una forma de vida extremament activa. Normalment, els espiracles i els sacs pulmonars romanen tancats per uns músculs especials que només es relaxen quan el contingut de diòxid de carboni de la sang assoleix o excedeix un 5%. D’aquesta manera, la pèrdua d’aigua, que invariablement acompanya larespiració, es manté en uns mínims.
Molts escarabats i d’altres insectes del desert obtenen humitat mastegant matèria vegetal higroscòpica que ha absorbit la humitat atmosfèrica durant la nit. Les calitges i boires també proporcionen una font d’humitat durant la nit en algunes regions desèrtiques, sobretot al Namib i Atacama. Alguns artròpodes sense ales, com ara les paparres, els peixets de plata, les puces i algunes paneroles i larves d’escarabat, són capaços d’absorbir vapor d’aigua directament de l’aire insaturat. El mecanisme que permet això encara es desconeix. Excepte en paparres i en la panerola nord-americana del desert (Arenivaga investigator), casos en els quals hi ha implicades peces bucals, el recte sembla ser el lloc per on s’absorbeix l’aigua. El fenomen no ha estat demostrat entre els escarabats adults del desert de la família dels tenebriònids, possiblement perquè ja han desenvolupat uns índexs tan baixos de pèrdua d’aigua que no els cal absorbir-ne més d’aquesta manera.
Tant els escarabats com els escorpins del desert són extremament resistents a la pèrdua d’aigua i tenen unes temperatures letals encara més altes. Els escarabats es refugien tant de la calor com dels enemics a sota de les roques, pedres i detrits. No excavenel sòl compacte, però algunes espècies, com Lepidochora discoidalis del Namib, tenen forma de plat i s’encauen ràpidament en la sorra solta. Per contra, els escorpins de les famílies diplocèntrids, vejòvids, càctids i escorpínids normalment tenen unes pinces inusualment gruixudes i pesades que els ajuden a l’hora d’excavar caus (simplement fan de suport mentre excaven bàsicament amb les potes). La majoria de membres de la família butíds, en la qual les pinces són lleugeres, no s’encauen sinó que habiten sota el pedruscall i la virosta, però entre altres excepcions figuren Leiurus quinquestriatus, que s’estén per tot el Sàhara Occidental i el Pròxim Orient, i Parabuthus hunteri, del Sudan. Es distingeixen quatre estils de vida diferents amb les corresponents adaptacions morfològiques entre els escorpins del desert: els litòfils o habitants de les roques, adaptats a la vida en esquerdes i clivelles de roques, amb el cos pla i allargat i les pinces dels pedipalps corbades que els permeten d’agafar-se fort en superfícies abruptes; els psammòfils, amb llargues pinces per excavar en la sorra solta; els fossorials, que es passen gairebé tota la seva existència en caus, des de les entrades dels quals es mouen activament mentre busquen menjar amb el cos gran i uns pedipalps semblants als dels crancs, amb una gran força per a aixafar; i els errants, que també es mouen activament mentre busquen menjar, amb el cos llarg i prim i pedipalps.
Molts deserts són habitats per grans taràntules (migalomorfs) que viuen en caus. Com els escorpins, normalment no surten durant el dia, quan serien vulnerables enfront dels seus depredadors. Altres famílies d’aranyes ben representades en entorns àrids són principalment saltícids, licòsids i tomísids. Als deserts nord-americans, les taràntules es troben en caus o forats de conills, mentre que les aranyes amb nius operculats i les taràntules excavadores constitueixen elements importants de la fauna del desert australià. Sembla probable que el predomini de les aranyes migalomorfes a Austràlia pugui ser provocat per l’absència de competència per part dels solífugs, ja que no n’hi ha. A l’Amèrica del Nord, també, els solífugs són menys dominants que en el gran desert paleàrtic, on les migalomorfes no són significatives. Les aranyes que habiten al desert es distribueixen a grans trets en dos grups ecològics: les grans i de vida llarga, que inclouen les migalomorfes excavadores i algunes de les licòsides més grans que també viuen en caus, i les petites, amb una vida comparativament més curta i nòmades, que inclouen les aranyes caçadores de les famílies gnafòsids, saltícids i tomísids. Les principals adaptacions de les aranyes al desert es basen en els hàbits d’encauar-se i en la compensació metabòlica per a l’alta temperatura.
La pugna entre predadors i depredats
A causa de la seva topografia plana i erosionada, com també de l’escassetat o l’absència de vegetació, els deserts són un dels medis terrestres en què els habitants estan més exposats als depredadors. Al mateix temps, als depredadors sovint els resulta difícil aguaitar la seva presa sense ésser detectats en un camp tan obert. Els depredadors capturen i maten les seves preses de diferents maneres i utilitzant una gran varietat d’armes (el fibló de l’escorpí, la teranyina de l’aranya, el camuflament i el verí del cròtal banyut, o el vol i el picat del falcó).
La majoria de predadors desertícoles cacen sense pauta fixa, movent-se aparentment a l’atzar fins que detecten la seva presa, tot i que alguns freqüenten indrets especialment favorables. Els carnívors del desert tenen dietes variades i poden desenvolupar mètodes específics per a enfrontar-se amb preses difícils. Per exemple, els icnèumons llancen els ous o els centpeus contra les roques per esclafar-los o els ratolins llagosters meridionals (Onychomys torridus) arrenquen d’una dentegada les cues dels escorpins abans de menjar-se’ls. En general, els carnívors s’alimenten d’herbívors, o d’altres carnívors, que són més petits o aproximadament de la mateixa mida que ells. Tot i que els lleons i els lleopards són capaços de matar animals molt més pesats que ells mateixos, la seva presa normal rarament ultrapassa el doble del seu pes. Els limitats recursos alimentaris de les espècies que viuen al desert no sempre poden mantenir bandes de llops o de gossos salvatges, animals que sí que són capaços de matar, aplegats, preses molt més grosses que ells.
Les formes de mobilitat dels rèptils i el seu valor tàctic
Els rèptils del desert mostren una varietat de sistemes de locomoció que són utilitzats tant per a obtenir menjar com per a refugiar-se. Les adaptacions per a soscavar no són especialment marcades, malgrat que en els llangardaixos i les serps del desert que ‘neden’ a la sorra s’observa que sovint tenen el rostre punxegut o en forma de pala i els seus narius poden estar dirigits cap amunt i no cap endavant. Els ulls, els narius i la boca els poden tenir protegits per vàlvules o altres estructures per a evitar-hi l’entrada dels grans de sorra quan submergeixen el cap en la sorra. Tenen el cos cobert de petites escates que causen poca fricció, i les seves cames es poden reduir, o fins i tot desaparèixer completament, de manera que la locomoció es porta a terme exclusivament serpentejant. Els amfisbènids també són soterradors sense potes amb unes escates molt fines.
Com a mínim 26 espècies de 7 famílies diferents de saures han desenvolupat unes escates sortides i allargades a les puntes dels dits. La morfologia d’aquestes puntes varia segons el tipus de substrat que freqüenten: les espècies que circulen per la sorra de zones ventoses, normalment tenen puntes triangulars, sobresortints i còniques; mentre que les riberenques que circulen per l’aigua tendeixen a tenir-ne en forma de rectangles més o menys amples.
Les serps es poden moure de diferents maneres, la més comuna de les quals la locomoció serpentejant. En aquesta, la força propulsiva es deriva de l’empenta de les corbes del cos contra prominències del terreny com ara pedres, tiges i altres irregularitats. Un altre mètode de locomoció és el moviment en acordió, que també es basa en la pressió dels anells del cos contra les irregularitats de l’escorça. La locomoció lateral permet que les serps de cos curt, com ara la cerasta o escurçó banyut (Cerastes cerastes), l’escurçó del Namib (Bitis peringueyi), i el cròtal banyut (Crotalus cerastes), es moguin a una velocitat considerable damunt de les superfícies llises i sorrenques, impedint al mateix temps que la major part del cos es posi en contacte amb la calenta superfície del sòl. Finalment, els pitons (Python) i els grans escurçons són capaços de lliscar cap endavant amb una locomoció rectilínia i amb el cos estès en una línia quasi recta. Les amples escates ventrals s’aixequen, es tiren cap endavant, es recolzen a terra, i arrosseguen el cos de la serp.
Les fatigues dels depredadors
Alguns depredadors cerquen la presa activament; d’altres, més nombrosos, la sotgen, de vegades camuflats, i l’agafen per sorpresa. D’altres, com ara els voltors, no capturen mai preses vives, però secundàriament s’han adaptat a una manera de viure basada a aprofitar la carronya. Sovint s’utilitzen combinacions de tècniques de depredació.
La selecció natural, generada per la competència entre els depredadors, pel menjar, i la seva presa, per evitar que se la mengin, comporta una veritable cursa armamentística entre depredador i presa. Al mateix temps, fins i tot els depredadors més poderosos com ara els lleons i els lleopards són, en alguna etapa de la seva vida, preses potencials d’algun altre depredador. Com a conseqüència, les seves adaptacions representen un terme mitjà entre aconseguir l’efectivitat en la cacera i evitar la captura. Per exemple, els màntids són depredadors rapaços pel que fa als insectes però, a la vegada, ells són víctimes d’ocells insectívors.
Molts depredadors cacen sense norma fixa, movent-se aparentment a l’atzar fins que veuen, ensumen, senten, toquen o detecten d’una o altra manera la seva presa. Fins i tot grans caçadors especulatius sovint augmenten les possibilitats de trobar presa buscant-la en llocs especialment prometedors, com ara tolls o trossos de vegetació suculenta. Els ocells rapinyaires tendeixen a caçar en àrees específiques que són freqüentades per preses potencials. La recerca activa de la presa es dóna a tots els tàxons de depredadors de l’ecobioma del desert i depèn de la locomoció.
Els animals depredadors de les terres àrides inclouen artròpodes, sobretot aràcnids i insectes, rèptils, ocells i mamífers. Menys importants són els miriàpodes, que només es troben al límit del desert, i els amfibis, dels quals n’hi ha unes quantes espècies de desert. En general, els amfibis no estan gaire ben adaptats com a depredadors. Mentre les granotes i els gripaus adopten l’estratègia de sotjar, la recerca activa és més comuna entre les salamandres. La presa és capturada traient la llengua enfora. La salamandra tigrada (Ambystoma tigrinum) dels deserts nord-americans captura insectes colpejant-los amb la meitat posterior de la llengua, a la qual queden enganxats a causa de la viscosa secreció glandular. Les granotes i els gripaus, d’altra banda, emboliquen la presa amb la punta de la llengua, que després tornen a introduir a la boca.
Entre els depredadors que sotgen la presa, es troba el cròtal banyut (Crotalus cerastes). Aquesta serp de cascavell, quan roman ajaguda, parcialment enterrada en la sorra, és quasi invisible per a les rates cangur i els altres petits animals que constitueixen la seva presa. Al mateix temps, malgrat el seu perillós verí, és vulnerable als seus propis depredarors específics, com ara el correcamins (Geococcyx californianus). D’una manera similar, els escurçons africans (Cerastes), els escurçons bufadors (Bitis) i les cobres (Naja) són víctimes del secretari (Sagittarius serpentarius), per no parlar de les mangostes. Menys acusat que en les adaptacions dels animals de presa, però així i tot present, hi ha l’efecte de la selecció natural sobre els mateixos depredadors basada en una depredació eficient (selecció per velocitat o silenci, cripsi agressiva, etc.). Els animals salvatges estan adaptats de manera simultània a molts factors del seu entorn, i això complica una anàlisi precisa.
La majoria de rèptils de desert són carnívors, i s’alimenten d’una gran varietat de preses, que poden ser tant vertebrats com invertebrats, depenent de la disponibilitat i la mida, i les troben a través d’una recerca activa. Molts llangardaixos petits s’alimenten quasi exclusivament d’insectes, mentre que les espècies més grans mengen llangardaixos més petits. Els varans devoren ous sempre que en troben, se’ls empassen sencers i els aixafen amb la contracció dels muscles de l’esòfag. El varà del Nil (Varanus niloticus) és conegut col·loquialment al Sudan com l’‘enemic del cocodril’ a causa de la seva predilecció pels ous de cocodril. Alguns llangardaixos, com ara l’agama comuna (Agama agama), són omnívors, i s’alimenten de pètals de flors, herba, fulles mortes, trossos de cacauets i altres matèries vegetals, com també de formigues i tèrmits. Es tornen més carnívors a mesura que es fan grans i, eventualment, s’ha observat que els adults s’alimenten canibalísticament dels seus propis descendents. Els llangardaixos de cua escato-sa (Uromastyx) són igualment omnívors, però el seu vegetarianisme s’accentua quan es fan grans. Per contrast, el diable espinós australià (Moloch horridus) i el llangardaix banyut de musell curt (Phrynosoma platyrhinos) són tots dos depredadors específics de formigues. Unes criatures tan lentes com aquestes serien particularment vulnerables si s’haguessin de refiar de fugir per a escapar-se dels seus propis enemics. Sovint es dóna per fet que per tal d’explotar la seva presa amb eficiència, els predadors han de permetre que les poblacions de preses mantinguin un nivell que maximitzi el nombre de preses que es poden capturar sense disminuir-ne la població. En el cas del llangardaix banyut i les formigues segadores de les quals s’alimenta, una estratègia de recerca permet la maximització de la disponibilitat de preses a un termini de setmanes o mesos més que no pas per hora o per dia. El nombre de llangardaixos banyuts és regulat per la disponibilitat i la productivitat de la seva presa, que és extremament alta.
Els llangardaixos de cua de fuet o “whiptail” (Cnemidophorus) cerquen la presa ficant els seus llargs musells sota els branquillons i agafant l’aliment del substrat, empaitant preses detectades visualment, i enfilant-se a la vegetació d’on foragiten la presa. Aquesta combinació d’estratègies fa que consumeixin una gran varietat d’animals. Moltes preses potencials amb coloració aposemàtica no són bones. Això implica la capacitat dels llangardaixos de cua de fuet de reconèixer una gran gamma de colors; la visió té una importància vital per als llangardaixos actius durant el dia a l’hora de cercar presa, tot i que l’olor també els pot servir d’orientació.
Mentre que els llangardaixos petits generalment s’alimenten només d’insectes, les espècies més grans poden menjar-se llangardaixos més petits. De la mateixa manera, les serps joves i les serps d’espècies més petites s’alimenten d’insectes i aràcnids, en contrast amb les espècies més grans que mengen principalment o exclusivament vertebrats. Les serps de desert petites solen ser depredadores generalistes. Les serps Chionactis occipitalis dels deserts nord-americans, per exemple, com la major part dels altres habitants del desert, són nocturnes, i cerquen insectes i les seves crisàlides, aranyes, escorpins i centpeus al ras. Per contrast, l’escurçó del Namib (Bitis peringueyi) s’amaga sota la sorra i surt per engolir les seves víctimes (llangardaixos com Aporosaura anchietae) durant el dia. A diferència de la major part de les altres serps, els seus ulls són situats a dalt del cap de manera que pot estar quasi totalment enterrat i a la vegada tenir un ampli camp de visió. Predador a l’aguait, s’ajeu amb la major part del cap i el cos enterrats, en la sorra relativament fresca, mentre espera llangardaixos i altres petits vertebrats, com ara ocells.
L’art d’escapolir-se
L’efecte evolutiu de la selecció de la presa per part dels depredadors ha engendrat un nombre considerable d’acusades respostes adaptatives. Per exemple, hi ha animals del desert que tenen coloració críptica, de manera que són del mateix color que el seu entorn àrid i sorrenc, o bé són negres i conspicus. Hi ha evidències considerables per suggerir que la coloració críptica ha evolucionat com una defensa contra els enemics depredadors mentre que la cridanera coloració advertidora o aposemàtica s’adreça a potencials depredadors per advertir-los que els seus posseïdors són repugnants o bé que disposen de formidables armes defensives. Ni la cripsi ni la coloració aposemàtica són exclusives dels animals del desert, però com que el desert és generalment un camp tan obert, tots dos són portats als límits més extrems de l’eficiència evolutiva.
Als deserts, les espècies vulnerables solen ésser críptiques i poc vistents. No sols els seus lloms són de colors similars als del paisatge del desert on viuen, sinó que a més les seves superfícies ventrals són pàl·lides, cosa que elimina l’efecte d’ombra. Els grills i les llagostes, que tenen un cos força sòlid, sovint semblen pedres, mentre que els pregadéus, amb un cos llarg i estret, més aviat semblen branquillons. Un bon exemple d’aquesta semblança protectora, el proporciona el llangardaix banyut de cua rodona (Phrynosoma modestum) dels deserts americans. Des de lluny, aquest petit llangardaix es confon amb el seu entorn, però de prop, es podria confondre fàcilment amb una pedra. Molts insectes petits de desert imiten les formigues, les abelles o les vespes, que són protegides per fiblons verinosos. Normalment, els himenòpters del desert són de color totalment negre. Això els fa ser conspicus davant dels enemics potencials per tal de no ser atacats i ferits de manera accidental per possibles depredadors. En regions temperades i als tròpics humits, on hi ha abundànciade vegetació, les abelles i les vespes normalment llueixen una combinació de groc i negre. Aquesta és una coloració molt conspícua en un medi com aquest, però, al desert, sobre un fons sorrenc, el negre pur és encara més conspicu, sobretot per a animals més petits com ara els insectes.
La coloració aposemàtica negra no es limita a avisar d’un fibló verinós. També pot indicar l’existència d’un gust o una olor fastigosos. Les mofetes tenen unes glàndules anals que segreguen un líquid tan fètid que quasi no tenen enemics; i anuncien això amb el seu color negre i blanc. Els tenebriònids també tenen tan mal gust que pocs animals a part dels solífugs se’ls mengen. També, com calia esperar, són ben negres: com en el cas dels escarabèids, la coloració negra, d’avís, va associada amb uns teguments ben durs i una olor i un gust desagradables. Els escorpins no se’ls mengen gairebé mai llevat que faci mesos que passen gana, però alguns solífugs se’ls mengen de bon grat. Els tenebriònids americans del gènere Elodes adopten una posició amb el cap abaixat quan estan enfadats i ruixen el seu enemic amb una secreció pudenta que conté benzoquinones. Això irrita la pell, i s’ha demostrat que repel·leix una gran varietat de predadors de manera molt eficient. No obstant això, no hi ha cap defensa perfecta, i els ratolins llagosters meridionals aprenen a emparar-se d’aquests escarabats empenyent endavant l’extremitat de l’abdomen de manera que les substàncies químiques repugnants que descarrega vagin a parar a la sorra sense perill.
Cridar l’atenció es pot aconseguir amb sons d’advertiment o amb colors aposemàtics. Els porcs espins són estrictament nocturs, però la blancor de les seves pues, quan les aixequen en un ventall característic, els fan ben palesos en la foscor. A més a més, quan un enemic els segueix de prop, els porcs espins fan tant soroll com poden, fent vibrar les pues de la cua (especialment modificades de manera que formen un sonall) i fent uns grunys roncs i guturals, i desprenent una fetor intensa. Si amb tots aquests mecanismes d’avís no n’hi ha prou per a dissuadir l’enemic de la persecució, el porc espí es llença enrere com un llamp i dóna un cop ben fort al seu perseguidor amb el manat de pues de sobre la cua. Com que són punxegudes i es desprenen fàcilment, aquestes pues poden produir una lesió greu a un depredador que sigui incapaç de treure-se-les.
Utilitzen el xiulet com a senyal d’advertència diverses serps i els únics saures verinosos, el monstre de Gila (Heloderma suspectum) i l’heloderma mexicà (H. horridum). Mentre les serps de desert són invariablement críptiques, tant amb finalitat ofensiva com de defensa, el monstre de Gila i l’heloderma mexicà són negres i grocs. Poden passar desapercebuts sota l’ombra clapejada de la vegetació del desert, però els seus colors són clarament aposemàtics quan són al ras. Fins i tot quan un animal està extremament ben defensat per verins, toxines i altres mecanismes químics i mecànics, sempre és millor que no l’ataquin que no pas haver-se d’implicar en una lluita durant la qual, tot i que en surti victoriós, podria resultar ferit. Aquest és el principibàsic de l’aposematisme. A les serps verinoses, que no els manquen enemics, només produeixen sons aposemàtics (xiulets, vibracions de la cua, etc.) quan se senten amenaçades. A banda els seus verins, les serps del desert són criatures petites i delicades en comparació amb els verinosos saures que són forts, de complexió pesada i molt més grans.
El ritme de la vida animal
La majoria d’animals del desert sobreviuen evitant les adverses condicions d’aquest hàbitat. Molts insectes tenen cicles vitals curts i només són actius després de la pluja, mentre passen la resta de l’any en un estat de repòs resistent a la sequera, normalment en forma d’ou. Sovint, els animals de vida més llarga també mostren ritmes estacionals d’abundància i s’amaguen durant el llarg període desfavorable de l’any. Per altra banda, les espècies que són actives durant tot l’any normalment són nocturnes i s’amaguen en caus i recers durant tot el dia.
La diürnitat i la nocturnitat
Tanmateix, hi ha algunes excepcions i no falten algunes espècies diürnes, moltes de les quals esdevenen crepusculars, és a dir, principalment actives a l’alba i al capvespre, durant l’estació calorosa. S’hi poden trobar diferents espècies d’escarabats, com també la majoria de les de llangardaixos i ocells. Al desert del Namib, per exemple, dues espècies de tenebriònids, l’una blanca, Onymacris langi, i l’altra negra, Physosterna globosa, responen a les altes temperatures de la superfície del sòl utilitzant un ritme d’activitat bimodal que els permet defugir la intensa calor de les hores de migdia. O. langi es posa en activitat més tard al matí i es retira més d’hora a la tarda que no pas P. globosa. Possiblement, el seu color blanc reflecteix més la radiació solar durant el període més calent del dia. Moltes altres espècies de tenebriònids utilitzen un ritme d’activitat bimodal similar, mentre que d’altres, com les nocturnes del gènere Lepidochora, eviten totalment l’excessiva calor del dia. Així i tot, una de les espècies d’aquest gènere, L. discoidalis, emergeix a última hora de la tarda, quan el vent refresca prou la superfície de les dunes per a permetre la recerca de menjar sobre les restes d’herba portades pel vent. Algunes espècies sembla probable que hagin desenvolupat l’activitat diürna en resposta a la competència amb les formes nocturnes.
La majoria de rèptils del desert són actius durant el dia. Emergeixen a l’alba i s’escalfen al sol fins que la seva temperatura corporal ha assolit un nivell òptim que mantenen combinant el sol i l’ombra. Alguns es posen a les branques dels arbustos del desert quan s’escalfen massa, i allà es refresquen amb el vent. De manera similar, els “gemsbok” (Oryx gazella) escalen regularment les dunes gegantines de Sossusvlei, a Namíbia, per tal de beneficiar-se de la brisa durant la calor del dia. El llangardaix excavador de les dunes (Aporosaura anchietae) que viu a la mateixa regió, es troba a la suau sorra moguda pel vent a prop de les crestes de les dunes, on la seva mitjana de temperatura preferida, 30-40°C, es dóna només durant un curt període de temps cada dia, estona que ha d’aprofitar per a buscar menjar abans que la sorra es torni tan calenta que s’hagi de submergir cap a un entorn més fresc a sota de la superfície.
Gràcies al seu eficient mitjà de locomoció, els ocells del desert poden defugir fàcilment les condicions desfavorables del desert. Busquen recer a l’ombra de roques, arbres, arbustos o qualsevol cosa que en faci durant les hores caloroses, i s’alimenten a primera hora del matí i a última hora de la tarda. Els voltors i altres rapinyaries planen sense esforç a molta altura durant les hores més caloroses del dia, mentre que els enganyapastors i les òlibes són nocturns, igual que els mamífers més petits (jerbus, gerbils, rates cangur, etc.). Les llebres del desert es refugien entre la vegetació, irradiant calor per les seves llargues orelles, mentre que els esquirols de terra seuen a l’ombra de les seves pròpies cues.
La sincronització de circumstàncies favorables
Per tal de sobreviure a la natura, totes les plantes i tots els animals vius han d’estar adaptats, i interactuar amb tot l’ecosistema del qual formen part. Les activitats fisiològiques i etològiques d’un animal, controlades pels seus rellotges biològics i condicionades pels sincronitzadors ambientals estan per tant adaptades, des d’un punt de vista temporal, a l’ecosistema. Aquests rellotges, amb un període aproximat d’un any, es coneixen com a circanuals; els rellotges amb una periodicitat lunar són circalunars; mentre que els rellotges diaris s’anomenen circadiaris. Per exemple, la reproducció d’alguns ocells del desert es dóna per una combinació de dos factors: el seu ritme reproductiu estacional i l’estímul de la pluja. Sense això són incapaços de pondre els ous, ja que la posta és inhibida. El mateix passa en el cas dels gripaus d’esperons americans (Scaphiopus), el ritme estacional de reproducció dels quals, combinat amb les vibracions i els sons provocats pels trons i les fortes gotes de pluja quan toquen a terra, estimula la seva emergència dels profunds caus on estiven.
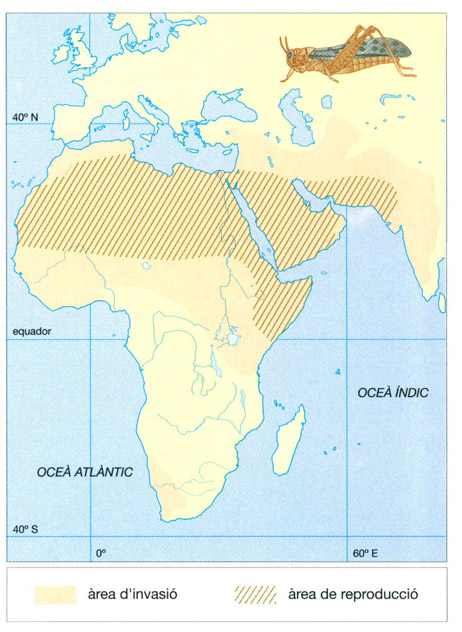
L’àrea de distribució de la llagosta pelegrina (Schistocerca gregaria) s’estén per les regions àrides d’Àfrica i el SW asiàtic, però s’han observat també petits eixams al N de la conca mediterrània, i fins i tot s’han trobat exemplars aïllats al S de les illes Britàniques. Mentre tenen prou menjar, les llagostes del desert es comporten exactament igual que qualsevol altra llagosta no migratòria. Quan l’aliment comença a escassejar, els individus en desenvolupament es converteixen en una forma gregària d’ales més llargues i colors marrons (la forma solitària és verda), que constitueixen eixams migratoris enormes. Aquests eixams devoren tot el que troben al seu pas.
IDEM, a partir de fonts diverses
Un altre exemple, una mica diferent, és el cas de la maduració dels òrgans reproductius de la llagosta pelegrina (Schistocerca gregaria), que és estimulada per terpenoides i altres substàncies químiques aromàtiques que desprenen estacionalment determinats arbustos del desert quan els brots foliars inicien la desclosa en el moment que comencen les pluges estacionals. Tot i que aquests compostos, responsables de les olors característiques de l’encens o de la mirra, s’han desenvolupat sens dubte com a repel·lents contra els insectes, pel que fa a la llagosta, més aviat caldria considerar-los afrodisíacs. Per aquests insectes, amb un cicle vital inferior a un any, però que fisiològicament han d’estar a punt per a aprofitar al màxim la pluja quan cau i estimula el creixement de gramínies i d’altres plantes anuals, tals compostos resulten d’allò més importants. El rellotge circanual de la planta regula amb ells indirectament el cicle reproductiu de les llagostes, de tal manera que aquestes són a punt per a la posta dels ous així que comença la pluja i per tant, en els moments previs al creixement de l’herba fresca i verda perquè els insectes acabats de sortir de l’ou s’alimentin.
Els rellotges circadiaris informen els animals del pas del dia a la nit o viceversa. Sense ells, els escarabats i escorpins de desert nocturns que viuen en profunds caus, no s’adonarien que ha arribat la nit, i per tant, el moment de sortir. Els rellotges biològics rarament són del tot precisos, però se sincronitzen diàriament amb el començament de la nit o de l’aurora. Els rellotges dels animals nocturns se solen sincronitzar amb el capvespre, i els de les espècies diürnes amb l’alba. Tot i que molts escorpins, com ara Paruroctonus mesaensis, són actius i emergeixen dels seus caus per caçar a la superfície del desert només una vegada cada quatre dies, els experiments de laboratori demostren que tenen els típics ritmes circadiaris. Presumiblement, aquests els fan pujar cada nit a la boca dels seus caus de manera que els seus rellotges se sincronitzen amb la intensitat lluminosa ambiental. El fenomen contrari es fa palès en el cas dels llangardaixos diürns.
A banda dels cicles de reproducció estacionals i els ritmes diaris de moviment i repòs, els rellotges biològics són importants pel que fa a la navegació que pren com a referència el Sol, la Lluna i els estels. La capacitat de guiar-se per la posició del Sol i d’analitzar el patró de la llum polaritzada a l’atmosfera permet a molts insectes i d’altres artròpodes, com ara la pastereta del desert Hemilepistus reaumuri, dirigir els seus moviments. D’una manera similar, els ocells migratoris, que travessen monòtones extensions de desert, orienten la direcció del seu vol observant la posició del Sol. Com que la direcció que prenen és la mateixa durant tot el dia, resulta que tenen en compte l’evident moviment del Sol en el cel entre l’alba i el capvespre. De manera similar, els ocells que volen de nit utilitzen la navegació lunar o bé es guien per les estrelles.
Des d’un punt de vista ecològic, els mecanismes de regulació interns tenen un paper important en el comportament i la fisiologia dels animals del desert. No obstant això, en la major part dels casos, el seu comportament és determinat pels estímuls rítmics ambientals, i els rellotges interns o endògens només els proporcionen una disponibilitat per a respondre a aquests estímuls de la manera apropiada en funció del temps. Així, són capaços de reaccionar a un estímul determinat al moment precís i no es deixen enganyar fàcilment per petites alteracions del seu medi. Malgrat tot, per importants que siguin els seus ritmes biològics, s’hauria de tenir present que la fisiologia i el comportament són determinats en primer lloc per respostes directes als estímuls ambientals i que aquests són modificats o accentuats en segon lloc pels rellotges biològics.
La fauna dels deserts sàharo-sindians
El Sàhara és molt vast i, des del punt de vista biogeogràfic, pertany a la vegada a la regió paleàrtica, que comprèn especialment Europa i una part d’Àsia, a la regió etiòpica, pròpia d’Àfrica, i a una regió sàharo-sindiana, que, a través dels deserts de Líbia, del Sinaí, de Síria, de Mesopotàmia i d’Aràbia, s’estén des de la costa atlàntica d’Àfrica fins al Pròxim Orient i l’Índia. És un desert molt antic i, al llarg de la història, les seves dimensions han variat molt a conseqüència de les modificacions de l’aridesa. Malgrat les afinitats d’una part considerable dels animals del Sàhara, sobretot els vertebrats, amb la fauna paleàrtica i etiòpica, la majoria dels que viuen als altres deserts sàharo-sindians de l’Àsia sud-occidental tenen un estret parentiu amb els saharians, de manera que és possible tractar conjuntament la seva fauna i la del Sàhara.
L’estoc faunístic saharià
Al llarg dels períodes més humits, algunes regions dels deserts que avui es coneixen eren verdejants i les creuaven rius, de vegades prou importants. En particular, la costa atlàntica del Sàhara occidental fou una zona de pas que permeté la invasió de les planes i les muntanyes de l’Àfrica septentrional per la fauna de les sabanes tropicals, al seu torn en continuïtat amb la de l’Àfrica meridional. Amb el temps, aquests intercanvis i moviments de la fauna, tant dins com al voltant del Sàhara, han donat a alguns animals la possibilitat d’adquirir les adaptacions necessàries per a viure en les condicions d’un desert càlid, de les quals les del Sàhara són el veritable paradigma, i per a esdevenir veritablement saharians. L’antiguitat del desert saharià, per altra banda, malgrat períodes humits que, de vegades, n’han reduït molt la superfície sense fer-la desaparèixer, ha permès l’evolució progressiva d’una sèrie d’espècies endèmiques, pertanyents a gèneres o a famílies originàries d’una o altra de les regions biogeogràfiques veïnes, que presenten adaptacions extremes a tots els problemes que planteja el medi desèrtic.
La major part d’aquestes adaptacions estan vinculades, directament o indirecta, a la limitació d’aigua utilitzable pels vegetals i, per tant, de recursos alimentaris per als animals. Aquests, tanmateix, s’adapten prou bé a les restriccions d’aigua, economitzant-la al màxim, amagant-se sota terra o en llocs protegits, on quasi sempre viuen en atmosfera saturada d’humitat i, eventualment, entrant en letargia. Però no poden viure sense menjar; per gran que sigui la seva frugalitat, els que poden alimentar-se prou són poc nombrosos i quasi sempre dispersos en els diferents medis que ofereix la immensitat del desert. Molts saben desplaçar-se a distàncies considerables i tornar al punt de partida o, quan convé, són capaços d’emetre senyals que poden ser percebuts per una parella allunyada, i facilitar així l’encontre que faci possible la perennitat de l’espècie. També els predadors estan adaptats a la frugalitat i disposen d’especialitzacions que els ajuden a localitzar les preses, els amenaça la mort d’inanició si esdevenen més nombrosos del que permet el nombre de preses disponibles. La predació ha tingut, certament, un paper important perquè limita encara més el nombre d’individus de les espècies-presa però tanmateix en permet la supervivència. Tampoc no s’ha d’oblidar els animals, en particular vertebrats, no pròpiament desèrtics, arribats al desert durant un període humit i atrapats pel retorn de la sequera en petits illots refugi no realment desèrtic; hi han romàs, d’alguna manera, com empresonats i sense experimentar noves adaptacions.
A primer cop d’ull, els paisatges saharians semblen pobres en vegetació i, encara que algunes plantes donen prova d’aptituds, sovint extraordinàries, per a créixer i multiplicar-se aprofitant la pluja més escadussera, la producció vegetal és notablement baixa i no permet alimentar massa consumidors primaris. Aquests consumidors primaris, al seu torn, ofereixen una biomassa alimentària limitada als consumidors secundaris. La irregularitat de les precipitacions i llur feblesa, per altra banda, fan especialment aleatòria la supervivència dels animals de vida aquàtica, en particular dels peixos i dels amfibis.
La important presència reptiliana
Els rèptils, a excepció de la família dels gecònids i d’algunes serps nocturnes molt petites de la família dels leptotiflòpids, suporten temperatures altes. Els que són carnívors troben l’aigua que necessiten en les preses de què s’alimenten, i els que són herbívors, en les plantes. Els més ben adaptats al desert necessiten temperatures especialment altes per a poder alimentar-se i desenvolupar una activitat normal. La major part volen una forta insolació i gairebé sempre s’exposen al sol. Al Sàhara, òbviament, predominen sobre els altres vertebrats.
La tortuga terrestre Testudo sulcata, gran i herbívora, força comuna a les sabanes meridionals, s’ha trobat fins al Ténéré. Al Sàhara, durant l’estiu, a manca d’aliment fresc, s’esmuny dins un cau que excava ella mateixa i hi roman inactiva. Una tortuga aquàtica i carnívora, la tortuga d’aigua ibèrica (Mauremys caspica leprosa), l’àrea de la qual s’estén al N fins als peus dels Pirineus orientals, s’alimenta de peixos a les basses del Sàhara.
La dels agàmids és una de les famílies de llangardaixos més ben adaptada a la vida al desert. Només es troba al Vell Món (Àsia i Àfrica) i a Austràlia. L’agama del desert (Agama mutabilis) és activa de dia i a l’estiu. A la nit s’arrecera sota una pedra. De dia li agrada situar-se en llocs elevats. L’agama de Bibron (A. bibronii) prefereix els penya-segats i les roques. L’agama de Tourneville (A. sarignii) s’enfila als cimals de les mates a les dunes, fins a Palestina. L’agama del Sinaí (A. sinaitica) viu a les dues ribes de la mar Roja. L’agama comuna (A. agama) de les regions de sabana més meridionals viu, al desert, a l’empara de tarteres rocalloses. El dob (Uromastyx acanthinurus), un agàmid gros, herbívor i insectívor, pesa prop de 500 g. Una espècie veïna, U. geyri, viu a les muntanyes del Sàhara central. Una altra espècie, el dob egipci (U. aegypticus) es distribueix a l’Egipte septentrional, a Palestina i al desert sirià.
La família dels gecònids és representada per una vintena d’espècies saharianes, insectívores i nocturnes, que viuen a les anfractuositats de roques o a les cases, on es desplacen per les parets o els sostres. Dues espècies mediterrànies, el dragó anellat (Tarentola annularis), pròpia dels arbres i de les roques, i el dragó comú (T. mauritanica) més antropòfila, i dues espècies d’afinitats tropicals, una de l’Ahaggar, T. ephippiata i l’altra de l’Àfrica occidental, T. neglecta viuen al Sàhara. A les regions rocalloses del Sàhara central i d’Aràbia, de vegades, se senten els crits sonors del gran dragó de Hasselquist (Ptyodactylus hasselquistii). Els petits dragons de dits fins, com el dragó pigmeu de Steudner (Tropiocolotes steudneri), el dragó pigmeu de Tripolitània (T. tripolitanus) o el dragó elegant (Stenodactylus stenodactylus) es distribueixen fins al N d’Aràbia on s’arreceren sota les pedres o a la sorra. Deixen les petjades de les potes al voltant de tota l’entrada del cau. El petit dragó de casc banyut (Geckonia chazalae) del Sàhara atlàntic s’arrecera sota les eufòrbies cactiformes. El dragó de pell rugosa (Cyrtopodium scaber) viu d’Egipte a la península Aràbiga i a l’Índia.
Al Sàhara, la família dels lacèrtids és representada pels veritables llangardaixos i sargantanes dels gèneres Acanthodactylus i Mesalina. La sargantana lleopard (A. pardalis) viu al Sàhara septentrional, a les zones més planes i amb menys sorra, de les ribes de les basses petites. La sargantana rugosa (A. boskianus), més gran, i la sargantana daurada (A. scutellatus) viuen preferentment a la sorra, en àrees localitzades però distribuïdes per tot el Sàhara i fins a la riba oriental de la mar Roja. Les mesalines tenen els dits menys llargs que les sargantanes del gènere Acanthodactylus. La mesalina clapada (Mesalina guttulata) viu en el límit septentrional del Sàhara com també al Pròxim Orient, i fins al desert de Sind. La mesalina de taques vermelles (M. rubropunctata), més gran, és més comuna.
La distribució de la família dels escíndids cobreix la part més gran del Vell Món. Algunes espècies presenten una reducció dels membres anteriors i posteriors i un cos fusiforme i llis. L’escinc o peix de sorra (Scincus scincus) es desplaça com un peix per la sorra de les dunes buscant les larves de què s’alimenta. La bívia ocel·lada (Chalcides ocellatus) té els membres anteriors més reduïts que els posteriors. La seva distribució s’estén cap a l’E fins al desert de Sind. La bívia de Schneider (Eumeces schneideri) i la bívia ratllada occidental (Mabuya vittata) viuen al Sàhara septentrional, el primer arriba a l’Afganistan i el segon fins al Pròxim Orient. La família dels varànids és representada pel varà del desert o varà gris (Varanus griseus) que pot abastar 1,5 m de llargada i la seva distribució s’estén cap a l’E fins a arribar a l’Índia.
Les serps que viuen al Sàhara tenen diferents estratègies per a alimentar-se. Els leptotiflòpids són serps molt petites i molt fines (per això s’anomenen de vegades ‘serps fil’), excavadores i que s’alimenten de petits invertebrats: Leptotyphlops macrorhynchus, per exemple, es distribueixen des del Sàhara fins al Sind. Pel que fa als colúbrids, els àglifs no tenen dents verinoses, aferren les preses entre les mandíbules i se les empassen tot seguit. La serp d’aigua (Natrix maura), comuna a l’Europa sud-occidental, viu també a l’Àfrica septentrional i arriba al Sàhara, on es pot veure, de vegades, a les basses més grans riques en peixos. Els colúbrids opistòglifs tenen ganxos verinosos al paladar posterior i enverinen les preses que s’empassen. La serp verda o colobra de Montpeller (Malpolon monspessulanus) pot fer 2,50 m de llarg i quasi no entra al desert; mentre que la colobra mallada (M. moilensis) es distribueix al voltant del Sàhara i a la península Aràbiga. La colobra diadema (Spalerosophis diadema) és sàharo-sindiana, com la colobra d’esquena vermella (Coluber rhodorachis). La colobra algeriana (C. florentulus) viu al límit septentrional del Sàhara i a l’Ahaggar. Els colúbrids proteròglifs enverinen llurs preses mossegant-les amb unes dents verinoses fixades al davant del maxil·lar superior. És el cas de la cobra egípcia (Naja haje), la distribució de la qual s’estén per les sabanes tropicals, davalla per la vall del Nil, i arriba fins als confins septentrionals del Sàhara i dels deserts d’Aràbia.
En els vipèrids, els ganxos verinosos, enormes i articulats, estan preparats per a inocular el verí des que la serp obre la boca i agafa la presa. L’escurçó banyut (Cerastes cerastes) viu en tots els deserts sàharo-sindians. L’escurçó saharià (C. vipera) més petit i sense banyes, només viu en la sorra del Sàhara i d’Aràbia. L’escurçó llevantí (Vipera lebetina) es troba sobretot al Sàhara septentrional, però també viu a Turquia, a Mesopotàmia i a l’Iran. A Egipte és substituïda per l’escurçó de les piràmides (Echis pyramidum).
Els ocells nidificants i de pas
Encara que hi hagi molts ocells de pas durant llurs migracions, no són nombrosos els que nien a les extensions desèrtiques que se succeeixen d’W a E de l’Atlàntic fins a l’Índia. Per això les àrees de nidificació sovint apareixen disjuntes, a causa de la competència local per l’aliment amb les espècies migradores.
L’ànec canyella (Tadorna ferruginea) fa niu a les roques, prop de l’aigua, al Sàhara nord-occidental, a Síria i a Turquia i de l’Iran a l’Afganistan. El voltor comú (Gyps fulvus), que, en canvi, no és saharià, també té aquesta distribució oriental. L’aligot gros (Buteo rufinus cirtensis) i el falcó llaner (Falco biarmicus) que poden considerar-se saharians, també tenen aquesta distribució oriental. L’hubara (Chlamydotis undulata) fa el niu al Sàhara septentrional, a la península Aràbiga, a l’Iran, a l’Afganistan, i encara més a l’E, a tots els deserts orientals. El torlit (Burhinus oedicnemus) es distribueix des del Sàhara central i septentrional fins al desert de Sind, igual que el corredor (Cursorius cursor). La ganga clapada (Pterocles senegallus) i la ganga coronada (P. coronatus) són pertot arreu, en petits grups, al Sàhara i fins al Balutxistan. El gran duc del desert (Bubo ascalaphus) i formes afins o formes locals del duc comú (B. bubo) estan repartides des del Sàhara fins al Pròxim Orient.
La terrerola sahariana (Ammomanes deserti), distribuïda des del Sàhara occidental fins al Balutxistan, i la terrerola de cua negra (A. cincturus), que es troba fins al Sinaí, l’Iran oriental i l’Afganistan són els alàudids més ben adaptats a la vida del desert. L’alosa puput grossa (Alaemon alaudipes) té la mateixa distribució, com també la carbonera del desert (Hirundo obsoleta) o el corb bru (Corvus ruficollis), la distribució del qual s’estén cap al NE fins al Sind. Entre d’altres còlits, el còlit del desert (Oenanthe deserti) té una distribució clarament sàharo-sindiana; el còlit de cap blanc (O. leucopyga) és un ocell endèmic saharià que també fa niu al Sinaí i a l’Aràbia; el còlit de Hume (O. alboniger), que no viu al Sàhara, fa niu des de l’E de l’Iraq i d’Aràbia, al S de l’Iran fins al Balutxistan i al Sind. El cuaenlairat (Cercotrichas galactotes) és nidificant al Sàhara central i a tota la regió mediterrània fins al Tadjikistan. Passa el mateix amb el barbollaire groc (Turdoides fulvus). La busqueta pàl·lida (Hippolais pallida) viu en els tamarius en tota la regió sàharo-sindiana. La boscarla del desert (Scotocerca inquieta) fa el niu al N del Sàhara, com també a Palestina, Jordània, Iran, i l’Afganistan. El tallarol saharià (Sylvia nana) té una àrea disjunta entre el Sàhara centroccidental i l’Iran oriental i l’Afganistan occidental. El botxí (Lanius excubitor) s’ha vist fent niu a tot el Sàhara, a Palestina i a l’Afganistan. El pinsà trompeter (Rhodopechys githaginea), l’hortolà (Emberiza striolata), i el pardal blanc (Passer simplex) són sàharo-sindians; però l’últim només viu a Àsia, a les sorres de l’E de l’Iran.
El cas dels mamífers
Els mamífers que viuen al Sàhara pertanyen, uns a la fauna paleàrtica, i altres a la fauna etiòpica. Alguns mamífers, però, s’han especialitzat tant a conseqüència de la llarga evolució que els ha conduït a viure en els deserts sàharo-sindians, que han esdevingut veritables endemismes que han anat colonitzant els espais desèrtics un darrere l’altre. Per això ha calgut que s’adaptessin a recol·lectar un aliment escàs escampat en extensions immenses en les quals formigues i ocells granívors els fan la competència. És el cas de la majoria de les espècies de rosegadors, dels eriçons o de les llebres, com també dels seus predadors, el nombre dels quals és, de mitjana, com arreu, proporcional a la quantitat d’aliment disponible. Alguns d’aquests animals, comparats als seus homòlegs de regions no desèrtiques tenen, de vegades, una densitat de població increïblement baixa, de tal manera que cada individu es pot trobar a molts quilòmetres d’un altre representant de la mateixa espècie. Alguns treballs realitzats sobre els rosegadors han posat de manifest correlacions entre la dispersió dels individus, l’aptitud per a tornar al cau des de distàncies llargues, la de conservar durant més temps el record dels llocs que han recorregut, i la hipertròfia de les ampul·les timpàniques. L’ampul·la timpànica és una expansió de l’orella mitjana i és lògic pensar que aquesta hipertròfia ha de tenir un paper en l’audició. En efecte, fins a un cert límit, el creixement en volum de l’ampul·la, que està plena d’aire, permet una millora de la transmissió dels sons de freqüència baixa.
Els mamífers, en una o altra forma, emeten sorolls diversos que corresponen a sons d’aquesta mena i són capaços de memoritzar la localització d’aquests sorolls, com també l’ambient sonor on viuen. Diversos rosegadors del desert saharià piquen el terra amb les potes posteriors, d’una manera rítmica, pròpia de cada espècie. Aquests senyals sonors, emesos per un rosegador a l’interior del seu cau, en certes condicions d’excitació, i especialment durant l’estació reproductora, són audibles pels humans a centenars de metres. Així, els mamífers del desert d’ampul·les hipertrofiades, insectívors, rosegadors, carnívors o fins i tot ungulats, tenen una sensibilitat més acusada pels sons, picades de peu o crits emesos per animals de la mateixa espècie. Aquesta hipertròfia és, per tant, una adaptació important adquirida després de llargs processos de selecció natural, a causa de la pressió de les necessitats reproductores, per tal com estan separats per llargues distàncies els uns dels altres i no poden deixar a l’atzar la possibilitat de trobar-se. En tot cas, sembla improbable que la predació hagi tingut un paper important en la selecció natural d’aquesta característica.
Al Sàhara, els insectívors només són representats per l’eriçó del desert africà (Paraechinus aethiopicus). Al Pròxim Orient és substituït per l’eriçó del desert d’orelles grosses (Hemiechinus auritus). L’ordre dels quiròpters només té tres espècies realment desèrtiques sàharo-sindianes. El rinopòmid de cua llarga (Rhinopoma hardwickei), el rat-penat orellut de Hemprich (Otonycteris hemprichi) i el rat-penat de nas triforcat (Asellia tridens). És en els rosegadors del desert que s’observen les especialitzacions més notables vinculades a aquest medi, en el qual tanmateix tenen una vida social i sexual ben activa. En els deserts sàharo-sindians només dues famílies de rosegadors han adquirit adaptacions tan eficaces que els permeten explotar conjuntament tots els medis del desert: els gerbíl·lids i els dipòdids. Al Sàhara s’hi afegeix la família dels ctenodactílids. En contrapartida, aquests rosegadors, si més no les seves formes més especialitzades, ja no poden recolonitzar els medis, més esponerosos, en els quals havien viscut els seus avantpassats llunyans; hi haurien de competir amb altres espècies, al seu torn incapaces de viure en les condicions de vida dels deserts, especialment sense aigua.
Els gerbíl·lids són representats, en els deserts sàharo-sindians, per cinc gèneres i nombroses espècies, totes principalment nocturnes menys la rata de sorra diürna (Psammomys obesus) del Sàhara i d’Aràbia i el gerbil gran (Rhombomys opimus) dels desertssindians (present també als deserts freds de l’Àsia mitjana i central). Totes les espècies es desplacen fent petits bots menys el gerbil de cua gruixuda (Pachyuromys duprasi) que camina i corre sense botar. El gènere Gerbillus comprèn moltes espècies de sola plantar nua. El gerbil nan (G. nanus) viu des del Sàhara Occidental fins a l’Índia. El gerbil campestre (G. campestris), una mica més gran que l’anterior, desborda el Sàhara cap al N, principalment el Marroc atlàntic. Del delta del Nil cap a l’E és substituïda pel G. dasyurus. Entre els gerbils de sola plantar peluda, el petit gerbil de la sorra (G. gerbillus) viu a les dunes, i el gran viu a les zones senzillament sorrenques per tot el Sàhara i Palestina. El gènere Meriones és representat per dues espècies, el jird libi (M. libycus) i el jird del desert (M. crassus), repartides per tot el Sàhara, Mesopotàmia, Aràbia, Iran i l’Afganistan. En les condicions normals del desert, totes dues espècies tenen una densitat de població molt baixa, de l’ordre d’un individu cada 10 ha, acompanyada d’una hipertròfia de les ampul·les timpàniques més gran. Totes dues espècies excaven caus complicats i profunds, en els quals el niu, com passa amb els de la majoria d’espècies de rosegadors del desert, es manté a la temperatura mitjana anual, propera als 23 o 25°C, en una atmosfera saturada d’humitat. El jird de Shaw (M. shawi) viu en el límit septentrional del Sàhara, sovint amb densitats de població elevades, de 50 a 100 individus/ha. En el seu règim no pot prescindir de l’aigua i només competeix amb el jird de Líbia als marges del desert. A partir de Palestina, aquesta espècie és substituïda per M. tristrami, les ampul·les timpàniques de la qual, com les del jird de Shaw, no estan hipertrofiades. Després, a l’Iraq i a l’Iran hi viuen principalment el jird pèrsic (M. persicus), i de l’Iran a l’Índia el jird del desert indi (M. hurrianae). La rata de sorradiürna (Psammomys obesus) viu en poblacions con-centrades, de 50 a 100 individus/ha, i les seves ampul·les timpàniques no estan hipertrofiades. És estrictament vegetariana; el seu règim consisteix en fulles i tiges que talla de les mates de quenopodiàcies que creixen sobre els terrenys salats. Consumeix cada dia el 80% del seu pes d’aquestes plantes, és a dir 2 g d’àcid oxàlic i més d’1 g de sodi, i excreta una orina quatre vegades més concentrada en sals que l’aigua de mar.
Al Sàhara, la família dels dipòdids és representada només per una espècie, el jerbu Jaculus jaculus. Encara que sigui més petit que el jerbu de l’Àfrica septentrional (J. orientalis), té ampul·les timpàniques molt hipertrofiades. Una sèrie de dipòdids dels gèneres Jaculus i Allactaga viuen en els deserts asiàtics, fins al Sind. La família africana dels ctenodactílids és representada pel gundí del Sàhara (Ctenodactylus vali), el gundí de M’zab (Massoutiera mzabi), i el gundí de Félou (Felovia vae). El gundí del Sàhara té ampul·les timpàniques més voluminoses que el gundí de l’Atles (Ctenodactylus gundi), la distribució del qual, pròpia de l’Àfrica septentrional, no arriba al desert.
L’ordre dels carnívors té, al Sàhara, quatre famílies: cànids, mustèlids, hiènids i felins. Els cànids sàharo-sindians són representats pel xacal comú (Canis aureus) i la guineu de Rueppell o de sorral (Vulpes rueppellii). El fennec (Fennecus zerda) viu al Sàhara i a Aràbia. La guineu de Blanford (V. cana) es distribueix principalment de Jordània al Balutxistan. A l’empara dels rius o dels uadis, la guineu comuna (V. vulpes), de distribució paleàrtica, penetra en el desert, on competeix amb la guineu de Rueppell, espècie sàharo-sindiana. Les ampul·les timpàniques dels exemplars de guineu comuna que viuen al Sàhara, i encara més les de la guineu de Rueppell i del fennec, estan hipertrofiades, i els pavellons de les seves orelles estan particularment desenvolupats. La família dels mustèlids, al Sàhara, és representada per una sola espècie, la mofeta de Líbia (Poecilictis libyca), les ampul·les timpàniques de la qual estan molt hipertrofiades. La hiena ratllada (Hyaena hyaena) és una espècie paleàrtica i sàharo-sindiana. El gat salvatge (Felis silvestris), un dels possibles avantpassats del gat domèstic, i el gat de sorral (F. margarita) són també dues espècies de felins paleàrtics i sàharo-sindians. Passa el mateix amb el caracal (F. caracal) i el guepard (Acinonyx jubatus), esdevinguts escassos al Sàhara.
Dins l’ordre dels hiracoïdeus, de la família dels procàvids es pot trobar el damà roquer (Procavia capensis), que viu a les zones rocalloses del S del Sàhara, a Palestina i Aràbia. L’ordre dels lagomorfs és representat per una forma desèrtica de la llebre del Cap (Lepus capensis) que es torna a trobar amb altres formes a tota l’Àfrica, com també a Europa i a Àsia. En un sol punt del Sàhara nord-occidental s’ha capturat la llebre de Whyte (L. whytei), una espècie repartida al S del Sàhara a tota l’Àfrica no sahariana.
De la família dels bòvids, dins l’ordre dels artiodàctils, al Sàhara es pot trobar l’addax (Addax nasomaculatus), la gasela dama (Gazella dama),la gasela comuna (G. dorcas), el “rhim” (G. leptoceros) i l’arruí (Ammotragus lervia). Al Pròxim Orient, la gasela persa o “djeiran” (G. subgutturosa) es distribueix des de l’Iraq fins al Balutxistan. Finalment, l’òrix blanc (Oryx leucoryx) també vivia a l’Iraq.
La família dels èquids, dins l’ordre dels perissodàctils, és representada pels ases salvatges asiàtics (Equus hemionus), de l’Iraq i Aràbia a l’E de l’Iran. L’hemió pròpiament dit (E. h. hemionus), a Síria i Mesopotàmia, avui extingit, i l’onagre a l’Iran (E. h. onager). Cal citar encara el camell comú (Camelus dromedarius), camèlid domèstic originari d’Aràbia, substituït pel camell bactrià (C. bactrianus) en els deserts freds d’Àsia.
La fauna dels deserts de Kalahari i del Namib
És del tot il·lusori, malgrat les seves formacions de dunes, voler definir una fauna de vertebrats pròpia del Kalahari i el Karoo. La major part de les espècies de mida mitjana o gran que s’hi fan són les mateixes que viuen a la sabana, habituades, arreu com aquí, a viure en condicions vinculades a pluges estacionals. El fet que un centenar de dies l’any, al desert del Namib, s’hi condensin boires matinals que no es dissipen fins ben avançat el matí, permet que, de la part més septentrional a la més meridional d’aquest desert, els llacs salats assecats, les extensions planes cobertes de còdols amb prou feines accidentades per alguns turons pedregosos i per les dunes que cobreixen més d’un terç de la superfície d’aquest desert, i finalment els sorrals siguin el marc de la vegetació i d’una multitud d’invertebrats. Alguns petits vertebrats obtenen també part d’aquesta aigua de condensació per a la qual han adquirit adaptacions particulars.
El cas dels rèptils i dels ocells
Entre les petites espècies de vertebrats que hi ha en aquests deserts, algunes són endèmiques, més aviat pròpies dels medis sorrencs. És el cas de les bívies cegues àpodes (Typhlosaurus) com la ratllada del Kalahari (T. lineatus), la de Gariep (T. gariepensis) o la de Brain (T. braini), com també de la bívia arborícola del Kalahari (Mabuya spilogaster) i de la bívia ratllada meridional del Namib (M. striata sparsa) que viu a les dunes d’aquest desert; ho és igualment del llangardaix de cua blava del Namib occidental (Cordylosaurus subtessellatus), del llan-gardaix ocel·lat del Namib (Meroles suborbitalis), del llangardaix excavador de les dunes (Aporosaura anchietae) o, encara, del dragó cridaner (Ptenopus garrulus), que se sent al Namib els vespres d’estiu, del dragó de terra del Kalahari (Colopus wahlbergii), del dragó de terra gegant del Namib (Chondrodactylus angulifer), del dragó diürn del Namib (Rhoptropus afer), com també de la diminuta serp nana de bec (Dipsina multimaculata) o de la serp de sorral del Namib (Psammophis leightoni) i de l’escurçó del Namib (Bitis peringueyi). Però força altres rèptils, molt menys especialitzats, també es troben ocasionalment en aquests deserts, tals com la bívia ratllada occidental (Mabuya occidentalis), l’escurçó Bitis caudalis, que també viu en contrades relativament humides, la cobra del Cap (Naja nivea), la tortuga del Cap (Pelomedusa subrufa), que aprofita la mínima bassa que troba al Kalahari, o la tortuga lleopard (Geochelone pardalis), que surt de la hivernada quan comencen les pluges d’estiu.
Entre els ocells cal citar la merla d’ales pàl·lides (Onychognathus nabouroup), el cua-roig del Kalahari (Cercotrichas paena), el tallarol formiguerbru (Myrmecocichla formicivora), l’abellerol de cua d’oreneta (Merops hirundinaceus), la guatlla arlequí (Coturnix delegorguei), el falcó pigmeu (Polihierax semitorquatus), el petit xot de cara blanca (Otus leucotis) i molts voltors i àguiles de vasta distribució.
L’àmplia diversitat de la mastofauna
Els mamífers grans són representats pels herbívors com el “gemsbok” (Oryx gazella), el nyu blau (Connochaetes taurinus), el búbal (Alcelaphus buselaphus), l’èland comú o èland del Cap (Taurotragus oryx) o el “springbok” (Antidorcas marsupialis) que, durant l’estació seca, sempre poden trobar aigua suficient en la sindriera (Citrullus lanatus) o en els cogombres de “gemsbok” (Acanthosicyos naudinianus) i cogombres africans (Cucumis africanus). També hi ha carnívors com el lleó (Panthera leo),la hiena tacada (Crocuta crocuta), la hiena bruna (Hyaena brunnea), el pròteles (Proteles cristatus), el guepard (Acinonyx jubatus), el xacal de llom negre (Canis mesomelas), la guineu del Cap (Vulpes chama) o la guineu orelluda (Otocyon megalotis).
Entre els mamífers més petits, es pot veure la lle-bre del Cap (Lepus capensis), la rata talp Cryptomys hottentotus, que s’alimenta dels cogombres de “gemsbok”, l’esquirol terrestre sud-africà (Xerus inauris), l’eriçó sud-africà (Atelerix frontalis), la suricata (Suricata suricatta) i força d’altres que poden sobreviure en aquest medi. Entre els rosegadors, algunes espècies de la família dels múrids com els ratolins ratllas sud-africans (Lemniscomys rosalia, Rhabdomys pumilio) han reeixit a sobreviure a Kalahari. La família dels gerbíl·lids, que al Sàhara ha donat les espècies més adaptades a viure en el desert, a Kalahari només és representada per un gerbil, Gerbillurus paeba, amb la sola plantar peluda i les ampul·les timpàniques relativament petites i per espècies que normalment viuen en les sabanes d’aquesta regió com ara Tatera leucogaster o T. brantsi. Al Namib es pot trobar el mateix Gerbillurus paeba, a més del gerbil de cua llarga (G. tytonis), amb les ampul·les més inflades, el de les dunes (G. vallinus), d’ampul·les molt inflades i cua llarga acabada en pinzell, i el de Setzer (G. setzeri), també amb les ampul·les molt inflades i la cua curta. També al Namib hi ha l’escàs gerbil de cua curta (Desmodillus auricularis) amb la cua curta i les ampul·les timpàniques particularment hipertrofiades. Es pot observar també la rata “dassie” o rata de tartera (Petromus typicus), que viu a les pedrusques com un damà, el ratolí de tartera nan (Petromyscus collinus), que circula entre les pedres com si fos un dragó gràcies als seus coixinets plantars i palmars adhesius, i el ratolí d’orelles grans (Malacothrix typica). L’únic mamífer que es coneix per no tenir homeotèrmia natural és el petit talp daurat de Grant (Eremitalpa granti), que circula per la sorra de les dunes del Namib, buscant invertebrats per menjar, ‘nedant’ com un peix dins l’aigua; s’ensorra a la profunditat on la temperatura de la sorra, que està en contacte amb el seu cos, li permet tenir una activitat òptima.
La fauna dels deserts càlids d’Amèrica del Nord
Aquests deserts no són tan àrids com el Sàhara, ja que les precipitacions tenen una mitjana de 130 mm i hi ha vegetació gairebé a tot arreu. Les seves faunes tenen poc en comú amb les dels deserts del Vell Món encara que són nombroses les espècies que comparteixen amb els biomes veïns, en els quals es troben elements tant holàrtics com neotropicals, i també amb les regions desèrtiques i subdesèrtiques d’Amèrica del Sud. Pel que fa als vertebrats, per exemple, la gran diversitat d’hàbitats que es poden trobar en el desert ofereix condicions acceptables per viure, almenys durant una part de la vida a molts d’ells encara que no estiguin específicament adaptats al bioma desèrtic. Altres, menys nombrosos, sí que estan adaptats a condicions on dominen la sequera i la dispersió dels recursos alimentaris característiques del desert.
La riquesa de l’herpetofauna
El rèptils són nombrosos. Entre els quelonis destaca la tortuga del desert Gopherus agassizii, d’hàbitat exclusivament desèrtic, que pot fer 37 cm, s’alimenta d’herba i de cactus i excava caus per fer-hi la posta. També són importants el dragó variegat (Coleonyx variegatus) que és nocturn i pot viure als deserts més secs arrecerant-se sota terra o sota les pedres i el dragó de dits plans Phyllodactylus xanti que grimpa sense parar per les roques.
La família dels iguànids comprèn, a més de les iguanes pròpiament dites, diferents ‘llangardaixos’ comparables als lacèrtids del Vell Món. La iguana del desert Dipsosaurus dorsalis viu en zones sorrenques; es mou entre les branques de les mates de les quals menja les fulles i les flors. La “chuckwalla” (Sauromalus obesus), viu entre les roques, a les esquerdes de les quals s’arrecera. Els umes (Uma notata, U. inornata, U. scoparia), iguànids amb els flancs marcats de negre i els dits vorejats d’escates espinoses, viuen a la sorra de les dunes i poden córrer redreçats sobre les potes posteriors. El gran llangardaix lleonat (Crotaphytus wislizenii), tacat de negre, viu a les planes àrides i semiàrides també pot córrer redreçat. Sceloporus magister, un dels llangardaixos espinosos del desert americà, té totes les escates punxants, s’alimenta d’insectes i de fulles pels turons de les regions àrides i semiàrides on viu, encara que alguns dels seus congèneres viuen a les selves del S de Mèxic; però el més abundant dels ‘llangardaixos’ de les regions àrides i subàrides de l’W nord-americà és el llangardaix de flancs tacats (Uta stansburiana), un petit iguànid marcat amb una taca negra darrere els membres anteriors. Un altre iguànid desèrtic és el llangardaix de cua llarga (Urosaurus graciosus); té una cua afilada que fa aproximadament dues vegades la llargada del cos. Entre els llangardaixos banyuts hi ha una espècie desertícola Phrynosoma platyrhinos; aquests petits iguànids de dieta especialitzada en formigues tenen una cua molt curta i són coberts d’escates espinoses, de les quals les situades darrere del cap són força grans. El llangardaix nocturn del desert (Xantusia vigilis) s’arrecera durant el dia sota les branques tombades al llarg de tota l’estació càlida.
La família dels teïids és principalment sud-americana, però a l’Amèrica del Nord té també 11 espècies d’una mena de sargantanes de llarga cua, anomenades “whiptails” (‘cues de fuet’) als Estats Units, tres de les quals viuen al desert (Cnemidophorus exsanguis, C. uniparens i C. tigris). La família dels helodèrmids és representada pel monstre de Gila (Heloderma suspectum), el cèlebre llangardaix verinós que viu a les vores de totes les zones àrides o semiàrides de l’Amèrica del Nord, on s’alimenta de petits mamífers i fins i tot de llangardaixos.
També el nombre de serps és considerable, des de les petites serps d’ulls vestigials (anomenades per això ‘serps cegues’) de la família dels leptotiflòpids, fins a la boa rosada. Les primeres, com per exemple Leptotyphlops dulcis o L. humilis, vermiformes i excavadores, prefereixen les “playas” de sòl sorrenc, en les quals s’enterren en recerca dels petits invertebrats que els serveixen d’aliment. La boa rosada (Lichanura trivirgata) és un bòid relativament petit (rarament supera 1 m de llarg) que busca més aviat indrets rocallosos i s’alimenta de petits mamífers i d’ocells.
Entre els colúbrids, hi ha serps nocturnes, com les de morro foliaci (Phyllorhynchus browni, P. decurttus), que viuen en els deserts sorrencs i són també excavadores, i també diürnes, com la ràpida Masticophis flagellum, una de les anomenades “whipsnakes” o ‘serps fuet’, o Arizona elegans, la serp lluent; una i altra eviten la vegetació i cacen a camp obert, encara que la primera freqüenta tota mena d’hàbitats i no es limita pas als deserts. També es troben al desert ocasionalment la serp lira californiana (Trimorphodon lambda), Salvadora hexalepis, amb el seu morro tacat, o Rhinocheilus lecontei, amb el seu llarg musell. Les serps de galons (Thamnophis), tant la de clatell negre (T. cyrtopsis) com la mexicana (T. eques), freqüenten els punts d’aigua. Moltes serps nocturnes excavadores, que utilitzen el nas rebaixat en la sorra com una pala (Chionactis occipitalis, C. palorastris, Chilomeniscus cinctus) són quasi exclusivament desèrtiques i s’alimenten de petits invertebrats. Les serps de cap negre (Tantilla), com la del desert (Tantilla planiceps transmontans) o la de Huachuca (T. wilcoxi), es distribueixen en petits claps de població discontinus.
La serp corall d’Arizona (Micruroides euryxan-thus) és el més destacat dels elàpids dels deserts càlids nord-americans, però les serps verinoses més conegudes pertanyen a la família dels vipèrids, són els cròtals o serps de cascavell. Els cròtals s’alimenten sobretot de petits mamífers i d’ocells i són coneguts pel sonall d’elements cornis que tenen a l’extremitat de la cua i fan sonar com a advertiment; molt difosos a diferents biomes americans, només dues espècies són quasi únicament desertícoles, el cròtal banyut (Crotalus cerastes) i el mojave (C. scutulatus).
Els ocells nidificants i de pas
Com a tots els deserts, s’observen molts ocells de pas. Només uns quants són sedentaris, entre els quals n’hi ha que viuen principalment a les muntanyes veïnes mentre d’altres són adaptats a les condicions de sequera i escassetat d’aliment pròpies dels deserts. L’ocell més gran de l’Amèrica del Nord és el còndor de Califòrnia (Gymnogyps californianus), que fins el 1870 era comú a les vores del desert; avui, però, només en queden algunes dotzenes d’individus, la majoria en zoològics. El xoriguer americà (Falco sparverius) és habitual a les planes i als deserts de Califòrnia, on nia als forats dels arbres o a les escletxes de les rocalles. La tórtora d’ales blanques (Zenaida asiatica), es pot veure en els oasis, on nia en colònies i visita els cactus quan els fruits són madurs. El correcamins (Geococcyx californianus) es troba als indrets àrids amb mates disperses; pot córrer a una velocitat de prop de 40 km/h i s’alimenta de grans insectes terrestres, de serps petites i de llangardaixos. El duc de Virgínia pàl·lid (Bubo virginianus pallescens) es manté en indrets del desert on creixen iuques de mida gran però generalment fa niu a les rocalles. El petit mussol follet (Micrathene whitneyi) viu en el desert de Colorado, allí on els cactus gegants han estat foradats pels picots per fer-hi caus. El mussol de llodriguera (Speotyto cunicularia) es troba en caus abandonats de petits mamífers del desert.
L’enganyapastors de Texas (Chordeiles acutipennis) descansa sobre el sòl cobert de grava del desert. L’enganyapastors de gorja blanca del desert (Phalaenoptilus nuttallii) entra en letargia durant l’hivern, quan no hi ha insectes. Un dels colibrís més petits, Calypte costae, viu als deserts càlids nord-americans i s’alimenta gràcies a les flors de iuca, de cactus i de diverses espècies de sàlvia. El picot daurat (Colaptes auratus) nia a l’W i al N dels deserts de Mojave i de Colorado, on fa forats als cactus gegants per instal·lar-hi el niu tal com fa també el picot de Gila (Melanerpes uropygialis). Al desert de Mojave encara es poden veure aloses banyudes (Eremophila alpestris), que es reconeixen pels plomalls negres disposats sobre el cap com si fossin orelles. El cargolet del cactus (Campylorhynchus brunneicapillus) si cal es refugia en les mates espinoses on instal·la el niu. El “sinsonte” o “thrasher” de Califòrnia (Toxostoma redivivum), és un ocell força comú que es pot observar buscant pel terra del desert els insectes, les aranyes i els fruits que li serveixen d’aliment i que instal·la el niu amagat a les maleses o entre les branques d’un mesquite. El papamosques de cua negra (Polioptila melanura), sedentari a les regions desèrtiques, on fa niu a les mates grans, s’assembla a un tallarol petit. El botxí de Sonora (Lanius ludovicianus sonorensis) captura les preses als altiplans desèrtics. El tallarol boscà de pit negre (Dendroica nigrescens) es manté en els terrenys ondulats. L’oriol de Scott (Icterus parisorum) viu a les iuques i als cactus, però s’alimenta d’insectes que caça al terra fora del desert. El passerell gola negre americà (Amphispiza bilineata) viu a les parts més assolellades del desert, on la vegetació és escassa.
El cas dels mamífers
Hi ha un cert nombre d’espècies de mamífers que es poden considerar endèmiques dels deserts càlids de l’Amèrica del Nord. Entre aquestes espècies, la musaranya Notiosorex crawfordi és pròpia de la Califòrnia meridional i del Mèxic septentrional. La guineu “kit” (Vulpes macrotis), característica per les seves orelles grans, l’extrem de la cua negra i la rapidesa de la seva cursa, es pot trobar a ple dia en els deserts de Mojave i de Colorado.
Els més nombrosos són els rosegadors, en particular les rates cangur (Dipodomis), els ratolins de bosses (Chaetodipus) i els esquirols de terra antílops (Ammospermophilus), així anomenats perquè sovint repleguen la cua sobre l’esquena tal com ho fa l’antílop americà (Antilocapra americana). Als deserts de Mojave i de Colorado es troben l’esquirol de terra antílop Ammospermophilus leucurus i l’esquirol de terra de cua de rata (Spermophilus tereticaudatus), mentre l’esquirol de terra de Mojave (S. mohavensis), més petit que el precedent, es pot trobar, quan no està en letargia, només al de Mojave. Els rosegadors més ben adaptats al desplaçament ràpid en els deserts americans són, però, sens dubte, les rates cangur, amb uns membres posteriors desenvolupats com els dels jerbus del Sàhara que els permeten desplaçar-se a salts; la rata cangur del desert (Dipodomys deserti) i la rata cangur de Merriam (D. merriami) destaquen entre les més freqüents. Pel que fa als ratolins de bosses, es caracteritzen per poder transportar importants provisions a les bosses que tenen a les galtes; el ratolí de bosses del desert (Chaetodipus penicillatus), el de Bailey (C. baileyi) i l’espinós (C. spinatus) estan restringits al desert més meridional, mentre que el ratolí de bosses de cua llarga (C. formosus) té la distribució desèrtica més vasta. La rata traginera americana del desert (Neotoma lepida) viu generalment associada amb els cactus, que li proporcionen els recursos d’aigua i al peu dels quals hi estableix el cau. El ratolí llagoster meridional (Onychomys torridus) viu als sorrals dels deserts de Mojave i de Sonora.
La fauna dels deserts càlids d’Amèrica del Sud
La fauna dels deserts de l’Amèrica del Sud és purament neotropical i, per això mateix, molt diferent de la de tots els altres deserts càlids, tret dels de l’Amèrica del Nord, que, com s’ha indicat, també han rebut tant la influència neotropical com l’holàrtica. Els ocells i els rèptils entre els vertebrats, i els artròpodes entre els invertebrats, són els grups més abundants.
El cas dels rèptils d’Atacama
El rèptil més característic de la costa desèrtica de Perú i Xile és l’iguànid Tropidurus peruvianus, un llangardaix robust, diürn, que es distribueix al llarg de la costa equatoriana, peruana i xilena, on se’n reconeixen nombroses subspècies, la més petita de les quals és precisament la que viu a Atacama, T. p. atacamensis; el seu aliment pràcticament es limi-ta a allò que la mar pot aportar: algues, petitscrustacis, restes de mol·luscs. A la part més meridional de la costa desèrtica hi ha un altre llangardaix, Callopistes maculatus atacamensis, de la família dels teïids, que coexisteix amb l’anterior i s’alimenta de la mateixa manera. Un iguànid insectívor, Liolaemus nigromaculatus bisignatus, que no sols s’alimenta d’insectes sinó també de petits amfípodes, viu a les dunes cobertes d’herba gelada o “rocio” (Mesembryanthemum crystallinum). Un dragó de dits plans, Phyllodactylus gerrophygus, s’alimenta, a la nit, d’insectes.
Als petits i esparsos claps de clavellines d’aire (la bromeliàcia Tillandsia humilis i altres congèneres) del desert de Tarapacá, una de les regions de clima més dur, en la qual, fora d’aquests claps, la vegetació és pràcticament inexistent, hi viu Phrynosaura reichei, un iguànid insectívor i ovípar, extremament rar. Més abundant resulta, fins al punt de ser dominant en aquest desert, Tropidurus tarapacensis, un altre iguànid, en aquest cas vegetarià i omnívor, que, com molts altres iguànids, es pot aixecar sobre les extremitats posteriors per córrer en posició bípeda. Amb ell viu també el petit dragó de dits plans Phyllodactylus gerrophygus. Les valls són també ocupades per formes de la costa com els ja esmentats Tropidurus peruvianus i Callopistes maculatus, acompanyats, més cap al N, de Liolaemus nigromaculatus i d’un dragó de dits plans més característic del Perú, Phyllodactylus inequalis, com també de serps, entre les quals destaquen Dromicus angustilineatus, que recorda la serp d’Esculapi (Elaphe longissima) europea, i Tachymenis peruviana, semblant a una colobra llisa (Coronella). Als oasis s’observa una iguana arborícola de petites dimensions, Tropidurus theresoides, i el ja esmentat dragó de dits plans, Phyllodactylus gerrophygus. A la regió supralitoral de l’Atacama, al peu de les pedres, hi viu una altra espècie de dragó, Garthia gaudichaudi.
Els ocells i els mamífers del desert costaner
Els ocells més característics de la costa desèrtica són grans carronyers, com el voltor negre americà o “zopilote” (Coragyps atratus), l’aura o voltor de cap roig (Cathartes aura) anomenat localment “jote”, que roba els ous i els pollets de les colònies d’ocells marins de les illes guaneres de la costa i el còndor dels Andes (Vultur gryphus), que s’alimenten de la carn dels cadàvers aportats per la mar. També assoleixen nivells d’abundància significatius altres rapinyaires com el falcó de Harris o “peuco” (Parabuteo unicinctus), el milà de cua blanca o “bailarín” (Elanus leucurus), l’àguila mora (Geranoaetus melanoleucus), els aligots (Buteo), el “tiuque” o “chimango” (Milvago chimango) o el xoriguer americà (Falco sparverius).
A les valls es poden observar el tirà vermelló (Pyrocephalus rubinus), l’“ani” (Crotophaga sulcirostris), l’alosa de pit vermell (Sturnella [= Pezites] bellicosa), la tórtora d’ales blanques (Zenaida asiatica) i el colibrí de Yarrell (Eulidia [= Myrtis] yarrellii). Mentre a camp obert apareixen els becadells granívors dits “perdicitas” (Thinocorus), el tinamú xilè o “perdiz” (Nothoprocta perdicaria), el “tapaculo” (Scelorchilus albicollis), la “tenca” o ocell burleta xilè (Mimus thenca), els tirans de terra o “dormilonas” (Muscisaxicola), els enganyapastors o “gallinas ciegas” (família dels caprimulgids), les “bandurrillas” (Upucerthia) o els “mineros” (Geositta), als rars espais humits es troben agrons o “garzas” (família dels ardeids), corriols o “chorlos” (família dels caràdrids) i els “canasteros” (Thripophaga). Diferents apòdids (falciots), hirun-dínids (orenetes), troglodítids (“chercantes”), muscicàpids (els papamosques coneguts localment com a “zorzales”), emberícids (els repicatalons i sits coneguts aquí com a “chirigües”, “chincoles” i “semilleros”) i fringíl·lids (els pinsans, passerells i similars, coneguts com a “jilgueros”, “diucas” i “yales”) inclosos els icterins o oriols americans, coneguts com a “loicas” i “tordos”.
En tota aquesta regió els mamífers són especialment escassos. Les colònies dels vampirs Desmodus depenen dels animals domèstics, mentre que diversos rats-penats insectívors viuen a costa dels insectes de les valls. Al llarg de la costa es troben otàries als punts favorables. Els escassos petits rosegadors, el pericot de Darwin (Phyllotis darwini) i diversos ratolins de camp americans del gènere Akodon viuen de la vegetació de la costa.
Els artròpodes més remarcables
Entre els artròpodes dominen els insectes i els aràcnids. Entre les famílies d’insectes més abundants i diverses figuren la dels proscòpids, els “palotes”, de l’ordre dels fasmòpters, i les dels acrídids, és a dir, les llagostes, i els tetigònids, és a dir, els saltamartins o “langostas cornudas”, de l’ordre dels ortòpters. De l’ordre dels coleòpters, els tenebriònids o escarabats negres, en particular els del gènere Gyriosomus, pul·lulen pertot arreu; també abunden els caràbids o escarabats terrers, els curculiònids o bequeruts, els melòids o escarabats butllofers, els estafilínids o cavallets i els melírids. Pel que fa als dípters, les famílies més ben representades són les dels agromízids o mosques minadores, els taquínids i els esciàrids, i pel que fa als himenòpters les dels formícids o formigues comunes, dels mutíl·lids o formigues de vellut i dels bracònids. De papallones o lepidòpters la família dels hespèrids (diürnes), la dels noctúids (nocturnes) i algunes de microlepidòpters assoleixen també nivells d’abundància considerable. Entre els aràcnids abunden sobretot els solífugs i els eritrèids o àcars vermells. En ocasions especials, aquests últims poden afegir al sòl, normalment de color marró grogós, un suau to vermellós.
La fauna dels deserts d’Austràlia
L’immens desert que s’estén sobre gran part del centre del continent australià és interromput de tant en tant per profundes valls al fons de les quals riberes i llacs permanents arreceren tota una fauna d’invertebrats i de peixos que se n’alimenten. Les temperatures poden superar els 40°C i les precipitacions són, en grans extensions, esporàdiques i febles, inferiors a 150 mm encara que, de vegades, les pluges torrencials transformen una part del desert en un immens llac temporal.
Una herba dura, l’herba porc espí (Triodia pungens), cobreix un terç de la superfície d’aquest desert i dóna refugi a tota una fauna de petits invertebrats, especialment formigues i petits insectes xucladors de saba. A la “mulga” (Acacia aneura) que creix per indrets, les formigues, més de 2 000 espècies de les quals són pròpies de les regions desèrtiques, es proveeixen d’una mena de mel. Les mates d’eucaliptus que creixen en els indrets més humits també aporten recursos alimentaris. Després de les pluges, les plantes efímeres, com els pèsols del desert (Clianthus), produeixen una gran quantitat de llavors ben nutritives. Però no hi ha fruits comparables als melons del desert de Kalahari o a les coloquintes del Sàhara, alhora alimentosos i plens d’aigua.
La singularitat de l’herpetofauna
Les nombroses granotes que apareixen després de la pluja es refugien sota terra durant els llargs períodes de sequera. Havent secretat un capoll mucós en el qual es tanquen, poden romandre en letargia durant molts mesos, fins i tot anys.
Els rèptils, remarcables per les seves adaptacions, són un bon testimoni de l’antiguitat dels deserts australians. A Austràlia hi ha una gran diversitat de llangardaixos, i n’hi ha que tenen nínxols ecològics que, en els altres continents, són ocupats per altres grups de vertebrats. En particular, els pigopòdids o llangardaixos serp (una família gairebé exclusivament australiana, amb algun representant escadusser en algunes illes veïnes) substitueixen les serps, els varànids fan el paper dels mamífers carnívors i alguns escíncids, i les petites bívies australianes, reemplacen els petits mamífers insectívors.
Les mates de “spinifex” tenen una funció en aquesta diversificació dels llangardaixos. Els protegeixen dels predadors i també són una reserva d’invertebrats entre els quals alguns troben les preses. La bívia Ctenotus piankai, el llangardaix serp Delma fraseri i el dragó Diplodactylus elderi estan constantment sota les mates. D’altres, com les bívies Ctenotus grandis i C. quattuordecimcostatus es mantenen al voltant. Finalment, n’hi ha d’altres que viuen en els espais nus, però sempre estan preparats per recuperar el recer d’una mata de “spinifex”. Entre aquests hi ha, sobretot, l’agàmid Amphibolurus isolepis i l’escíncid Ctenotus calurus.
En el desert australià hi ha més espècies de rèptils nocturns (dragons, bívies o llangardaixos serp que d’aquesta manera eviten la calor del dia) que en el desert de Kalahari o a l’Amèrica del Nord. Tanmateix, algunes espècies d’Amphibolurus i de Ctenotus són actives a ple dia, igual que la majoria dels llangardaixos del Sàhara.
L’endemicitat dels ocells
Austràlia té quasi 650 espècies d’ocells endèmics o propis d’aquest continent i de les regions veïnes, a les quals cal afegir 34 espècies introduïdes o de pas. Una part d’aquests ocells nidifiquen a tot Austràlia, compresos els deserts, si bé algunes espècies es limiten al conjunt de les regions situades al N dels 20°S, d’altres al de les situades al S dels 30°S, i finalment n’hi ha que fan niu entre aquestes dues latituds. Les espècies que es poden considerar aptes per a reproduir-se a les regions desèrtiques, atès que hi nidifiquen, no arriben a 90 i encara només algunes tenen una distribució limitada al desert.
D’altres tenen distribucions molt àmplies, que excedeixen de bon tros els límits del continent australià, com dos rapinyaires pràcticament cosmopolites: el falcó pelegrí (Falco peregrinus) i l’òliba (Tyto alba), i la piula grossa (Anthus novaeseelandiae). Comptat i debatut, només el cargolet australià fosc o “dusky grasswren” (Amytornis purnelli), de la família dels malúrids; el bec espí pissarrenc o“slaty-backed thornbill” (Acanthiza robustirostris), de la dels pardalòtids, i el menjamel de cap gris o “grey-headed honeyeater” (Lichenostomus keartlandi), de la dels melifàgids, són els únics ocells australians endèmics del desert central. Es tracta, és clar, d’ocells de famílies de la fauna australiana, amb prou feines amb alguna espècie difosa fins a Nova Zelanda o altres illes del Pacífic o fins a Insulíndia. A banda els rapinyaires esmentats, n’hi ha uns quants més, tots diürns, que s’alimenten de la fauna dels deserts i els subdeserts australians: l’àguila de cua en tascó (Aquila audax), l’àguila calçada australiana (Hieraaetus morphnoides), el berigora o falcó bru australià (Falco berigora) o el falcó comú australià (F. cenchroides).
La petita tórtora diamant (Geopelia cuneata), endèmica, és pròpia de l’W d’Austràlia i dels deserts centrals. El tudó crestat (Geophaps [=Ocyphaps] lophotes), també endèmic, viu a l’W i a l’E. En canvi, el tudó de “spinifex” (G. [=Petrophassa] plumifera), endèmic al NW, s’ha trobat niant al desert central. Els lloros i afins, que a Austràlia són nombrosos, nien majoritàriament a les zones arbrades. Tanmateix, la cacatua negra de cua vermella (Calyptorhynchus banksii [= C. magni-ficus]) es pot reproduir al centre del continent, com la cacatua de cap rosat o “galah” (Eolophus [=Cacatua] roseicapillus), la cacatua rosada o de Leadbeater (Cacatua leadbeateri), el periquito (Melopsittacus undulatus), la cotorra de Port Lincoln (Platycercus[= Barnardius] zonarius), del tot absent a l’E del país, o la cotorra de “mulga” o cotorra multicolor (Psephotus varius), que té la mateixa distribució geogràfica que la “mulga”, però de la qual, tanmateix, no depèn ni per a alimentar-se ni per a nidificar.
El cucut bronzejat de Horsfield (Chrysococcyx basalis) és l’únic cucut australià que nidifica al desert. L’enganyapastors australià (Podargus strigoides), el més gran dels caprimúlgids, només necessita un clot a terra per a fer-hi el niu on la femella pon els ous, de la incubació dels quals únicament s’ocupa el mascle. L’endèmic blauet d’esquena vermella (Todirhamphus [=Halcyon] pyrrhopygia) es troba pertot arreu, encara que suporta malament les grans pluges, que malmeten el seu niu, i prefereix els ambients menys plujosos mentre l’abellerol irisat australià (Merops ornatus) es reprodueix allà on troba els insectes dels quals s’alimenta.
L’oreneta d’esquena blanca australiana (Cheramoeca leucosternus) només nia al centre i S d’Austràlia, igual que l’oreneta arborícola o “tree martin” (Hirundo nigricans), la distribució de la qual desborda àmpliament els límits d’Austràlia; mentre que l’oreneta ariel o fairy martin (H. ariel) només és absent del Gran Desert de Sorra. El cucut botxí de cara negra (Coracina novaehollandiae) es reprodueix a tot Austràlia excepte al Gran Desert de Sorra, una distribució comparable amb la del refilador d’ales blanques (Lalage sueurii). El bocamoll coronat o “grey-crowned babbler” (Pomatostomus temporalis) fa niu més aviat al N, inclòs el desert central, mentre que el bocamoll de celles blanques o “white-browed babbler” (P. superciliosus) és propi del S del continent, com l’alosa australiana roja o “rufous songlark” (Cinclorhamphus mathewsi) i l’alosa australiana bruna o “brown songlark” (C. cruralis).
El cas dels mamífers
Els mamífers australians són principalment marsupials. Tanmateix, encara que arribats molt recentment a l’escala geològica, hi ha alguns placentaris que han tingut prou temps per a adquirir especialitzacions eficaces. És el cas d’alguns rosegadors de la família dels múrids, concretament de la subfamília dels hidròmids, que són a Austràlia des de fa només 5 o 10 milions d’anys. Així, entre les nou espècies de ratolí jerbu australià (Notomys), totes amb potes posteriors allargades i cua llarga amb una tofa de pèls llargs a l’extrem, només una, el ratolí cangur de “spinifex” (N. alexis), viu en el desert central d’Austràlia i hi està tan ben adaptat que pot prescindir d’aigua per beure. En el mateix medi viuen dues espècies de ratolins comuns australians (Pseudomys): el ratolí australià de sorral (P. hermannsburgensis), que és social, i el ratolí australià bru del desert (P. desertor), que, al contrari de les altres 18 espècies del mateix gènere, és principalment herbívor i solitari.
El cas dels dromedaris, introduïts en el continent australià com a animals de càrrega, és ben diferent del d’aquests ratolins. Tanmateix, esdevinguts inútils per a l’objecte per al qual havien estat introduïts, foren abandonats en gran nombre i s’assilvestraren amb tant d’èxit que, actualment, en viuen uns 20 000 al desert australià, cosa que representa, de fet, la població salvatge més nombrosa del món d’aquests animals.
Al desert australià hi ha representades quatre famílies de marsupials: la dels tilacòmids, la dels dasiúrids, la dels notoríctids i la dels macropòdids. Fins el 1930 hi vivia també un representant de la família dels peramèlids, el bàndicut de morro llarg del desert (Perameles eremiana), però des d’aquella data no s’ha tornat a trobar i es considera extingit. A la primera de les famílies esmentades pertany només el bàndicut orellut o “bilby” (Macrotis lagotis), un animal que pesa aproximadament 1,5 kg; excava caus profunds que li permeten fugir de la calor i menja sobretot tèrmits.
Els dasiúrids són representats al desert per moltes espècies, totes de musell punxegut i alimentació insectívora o carnívora; totes fan niu en un cau subterrani però els seus desplaçaments són per la superfície. El “kultarr” (Antechinomys laniger) és com una mena de ratolí marsupial solitari, amb la cua més llarga que el cos i les extremitats posteriors també molt llargues i dotades de 4 dits, que corre amb lleugeresa. El musell és molt punxegut, les orelles i els ulls grans i el pelatge llarg i llanós. La “mulgara” (Dasycercus cristicauda) viu a les dunes; és més gran i de pelatge més suau i dens que el del “kultarr”; és un animal social, d’orelles curtes, peus amb 5 dits i una cua fina i guarnida amb un pinzell de pèls negres a l’extrem; cada dia consumeix la quarta part del seu pes en insectes i així pot viure sense beure. El “kowari” o ratolí marsupial de Byrne (Dasyuroides byrnei) és el més gran dels ratolins marsupials dels deserts australians i pesa de 70 a 150 g; les orelles són curtes i la cua porta una mena de pinzell negre molt gruixut a l’extrem; en els deserts pedregosos i a les zones de vegetació esparsa és solitari i actiu de nit; pot arrecerar-se en un cau que ell mateix excava o en el d’un “bilby” i, quan fa fred, entrar en letargia. Molt més petits són els “ningauis” (Ningaui ridei i N. timealeyi), uns ratolins marsupials diminuts que només pesen de 2 a 12 g; tenen, com els anteriors, un musell punxegut, orelles arrodonides i una cua llarga i fina i els seus peus fan 2 mm d’amplada; són nocturns i solitaris. El primer, el més desèrtic de tots dos, té ampul·les timpàniques hipertrofiades com els rosegadors desèrtics del Sàhara, i pot entrar en letargia. Un altre dasiúrid insectívor és el ratolí marsupial de cua gruixuda (Pseudantechinus macdonnellensis), la cua del qual pot engruixir-se a la base quan el règim alimentari li permet acumular reserves de greix. El mateix passa en el cas d’alguns “dunnarts” (Sminthopsis), com el d’Ooldea (S. ooldea), descobert el 1970, el de cua gruixuda (S. crassicaudata), el de cara ratllada (S. macroura), i probablement també el de peus peluts (S. hirtipes), particularitat poc freqüent en els ratolins marsupials, versemblantment vinculada als desplaçaments damunt la sorra.
La família dels notoríctids només és coneguda pel talp marsupial (Notoryctes typhlops), que és relativament abundant a tots els sorrals del desert australià; ocupa el mateix nínxol ecològic que el que ocupa el talp daurat de Grant (Eremitalpa granti) als deserts de l’Àfrica austral. També es poden trobar als deserts australians alguns macropòdids, en particular el “wallaroo” comú o euro (Macropus robustus), comú a tot Austràlia, i el cangur roig (M. rufus), algunes poblacions del qual s’adapten perfectament a l’aridesa.