Els orígens i la diversitat de la flora dels deserts i els subdeserts càlids
Hom pot demanar-se si existeix, a escala global, una flora específica dels deserts i els subdeserts, i si hi ha uns determinats grups taxonòmics de plantes limitats a aquests ambients, però la realitat és que no. La flora dels deserts i els subdeserts és formada per elements certament especialitzats, però procedents de les grans regions florístiques on es troben.
La multiplicitat dels orígens
La flora mediterrània dóna el fonamental del poblament vegetal dels deserts i els subdeserts ‘mediterranis’ (Sàhara septentrional, erms subdesèrtics de l’Àfrica septentrional, erms i timonedes de la regió mediterrània). El resultat és una flora amb predomini de l’element mediterrani, tant en l’escala de la família (cariofil·làcies, cistàcies, crucíferes, geraniàcies, labiades o lamiàcies, umbel·líferes o apiàcies, ranunculàcies, escrofulariàcies) com fins i tot del gènere, amb diferenciació d’algunes espècies pròpies.
Els deserts tropicals del Vell Món són colonit-zats per una flora paleotropical, caracteritzada per certes famílies com acantàcies, aïzoàcies, apocinàcies, asclepiadàcies, bignoniàcies, bombacàcies, caparidàcies, celastràcies, cesalpiniàcies, cloridiàcies, combretàcies, commelinàcies, esterculiàcies, loganiàcies, malpighiàcies, moràcies, moringàcies, nictaginàcies, olacàcies, oleàcies, pedaliàcies, rubiàcies, salvadoràcies, sapindàcies, simarubàcies, zigofil·làcies, etc.; per subfamílies particulars (andropogoides, panicoidees) de famílies de distribució més àmplia o fins i tot cosmopolita (com, en aquests dos casos, la de les poàcies o gramínies), i per gèneres i espècies més o menys confinats a les zones intertropicals del Vell Món. Un cas particular seria el de la “rand flora”, la flora xeròfila dels deserts i subdeserts de l’Àfrica austral. La “rand flora” comprendria 1 140 gèneres, dels quals 420 endèmics, i 12 700 espècies, de les quals 10 500 són igualment endèmiques, entre elles 1 900 aïzoàcies, 1 800 compostes o asteràcies, 1 100 lleguminoses, 800 ericàcies, 800 liliàcies, 800 iridàcies i 500 asclepiadàcies.
Els deserts i subdeserts càlids del Nou Món presenten una flora neotropical, que es caracteritza per un cert nombre de famílies i de gèneres exclusius o quasi exclusius del Nou Món, tal com les agavàcies, cactàcies, bromeliàcies, fouquieràcies, garriàcies, hidrofil·làcies, krameriàcies, loasàcies, passifloràcies, taxodiàcies, turneràcies, etc.
Una excepció a aquesta multiplicitat d’orígens segons la localització dels diferents deserts la constituiria la flora halòfila. Els halòfits, a escala mundial, pertanyen a grups taxonòmics prou definits, encara que variats. Existeixen unes 6 000 espècies de fanerògames halòfiles terrestres i 2 000 espècies litorals (manglars) i pròpiament marines (hidròfits marins). Dels halòfits terrestres, un 25% són quenopodiàcies, mentre que un 10% són gramínies, un 4% compostes, un 4% més plumbaginàcies i la resta es reparteixen en un gran nombre de famílies, entre les quals destaquen aïzoàcies, ciperàcies, fabàcies o papilionàcies, tamaricàcies i zigofil·làcies.
La diversitat ignorada

Les palmeres són les plantes més representatives dels indrets humits de les zones àrides. En els diversos biomes, n’existeixen més de 2 500 espècies, repartides en uns 200 gèneres. El seu aspecte és inconfusible: una tija més o menys lignificada i amb aspecte de tronc, però sovint molt esvelta i rarament ramificada, coronada per un plomall de fulles grosses, palmatisectes o pinnatisectes i amb un llarg pecíol. Hi ha algunes espècies, però, que presenten un tronc ramificat dicotòmicament, com la palmera dum (Hyphaene thebaica), àmpliament distribuïda per bona part del continent africà. D’ecologia molt variada, les palmeres habiten des de les selves plujoses, on estan molt ben representades, fins a les sabanes i els deserts. Tanmateix, les seves necessitats d’aigua subterrània, ja sigui superficial o en profunditat, les concentren en aquest darrer bioma als oasis i a les valls fluvials. La seva àrea de distribució és molt àmplia, amb representants a tots els continents, bé que on estan més diversificades és a l’Àsia. Tanmateix, gairebé totes les espècies es limiten a una determinada regió, i molt poques tenen una distribució àmplia per més d’un continent, com el gènere Borassus, que es troba per l’Àfrica i l’Àsia. Altres espècies de distribució àmplia són les cultivades als oasis, essent la més coneguda la palmera datilera (Phoenix dactylifera) originària del delta del Tigris i l’Eufrates, on s’han descrit les primeres plantacions, datades del 4000 aC; actualment es cultiva a moltes altres regions tropicals i subtropicals àrides pels seus fruits, els dàtils, i per a moltes altres utilitats. Una altra palmera molt coneguda és la palmera de ventall californiana (Washingtonia filifera), cultivada com a ornamental en parcs, jardins i carrers.
Jordi Corbera
L’anàlisi global de la flora dels deserts i els subdeserts càlids està per fer. Només hi ha inventaris detallats per països i, de vegades, per regions, però aquests inventaris es realitzen generalment segons bases administratives més que no climàtiques o ecològiques. Cal basar-se, doncs, en alguns exemples precisos i ben documentats per a intentar una estimació global.
Uns bons termes de partida i de comparació poden ser les flores de les estepes subdesèrtiques del N de l’Àfrica i del Karoo de l’Àfrica austral. Segons una anàlisi recent molt detallada, els erms de les zones àrides de l’Àfrica septentrional (uns 630 000 km2), entre l’Atlàntic i la mar Roja, comprenen 2 600 espècies. La riquesa florística d’aquesta zona és, doncs, de 41 espècies per 10000 km2. El Karoo de l’Àfrica austral (500 000 km2) és una zona comparable des de nombrosos punts de vista; s’hi fan, però, prop de 7 000 espècies, o sigui, 140 espècies per 10 000 km2. El conjunt de la flora fanerogàmica de l’Àfrica septentrional s’estima en 8 000 espècies, mentre que la de l’Àfrica austral comporta 24 500 espècies; les zones àrides extra-tropicals africanes representen, així, el 33% de la flora total de l’Àfrica septentrional i el 29% de la flora austroafricana. Aquestes xifres subratllen el fet, prou conegut pels botànics i pels fitogeògrafs, de l’extraordinària riquesa de la flora sud-africana, 180 espècies per 10 000 km2 contra només 10 per l’Àfrica septentrional. Cal tenir en compte, però, que en aquesta regió la disminució de la xifra es produeix com a resultat de la important proporció de superfície sahariana; si no es té en compte la zona sahariana, la riquesa de la flora de l’Àfrica septentrional és de 65 espècies per 10 000 km2, és a dir, una riquesa florística que arriba a representar el 36% de la de l’Àfrica meridional.
El conjunt d’espècies dels deserts i els subdeserts càlids mundials es pot arribar a estimar aproximadament en unes 50 000 espècies, o sigui, del 15 al 20% de la flora fanerogàmica mundial (atès que les estimacions varien de 250 000 a 300 000 espècies), distribuïda sobre un territori que comprèn prop del 23% de la superfície de les terres del planeta. Aquesta xifra pot semblar elevada, però s’ha comprovat que, per exemple, les zones àrides de la conca mediterrània, molt estudiades, posseeixen més del 23% del total d’espècies de la flora mediterrània (aproximadament 22 000 espècies). Les zones seques africanes posseeixen prop de 18 000 espècies sobre un total continental de 68 500, o sigui, el 26%.
Els tipus biològics i les formes de la vida vegetal
En tot el regne vegetal, independentment de l’ecosistema i el règim climàtic, les plantes han desenvolupat adaptacions morfològiques, anatòmiques i fisiològiques per tal d’optimitzar la seva capacitat de sobreviure. Algunes espècies no emparentades, pertanyents a famílies diferents, han desenvolupat adaptacions convergents en resposta a pressions selectives similars. Enlloc és això tan evident com als durs ambients de deserts i subdeserts.
Els tipus biològics

Les diferents formes biològiques de les plantes responen a estratègies de supervivència també diferents, i reflecteixen les defenses que cada espècie vegetal ha adoptat contra les temperatures mínimes hivernals. Aquests cinc tipus, establerts pel botànic danès Raunkiaer (1860-1938), es basen en l’alçada respecte al nivell de terra a la que se situen les gemmes perdurants. Els faneròfits són principalment arbres i arbustos amb les gemmes elevades, a més de 25 cm de la superfície del sòl, i, per tant, totalment exposades al medi. El camèfits són arbustos més baixos, amb les gemmes també aèries però situades molt més a prop del sòl. Els hemicriptòfits estan constituïts per plantes herbàcies amb les gemmes al nivell de terra, i els criptòfits ja les tenen amagades sota el sòl (geòfits) o sota l’aigua (hidròfits). Finalment, els teròfits són plantes anuals que passen l’època desfavorable en forma de llavors. A les regions àrides les formes de vida vegetal més freqüents són els teròfits, que predominen a les zones amb pluges estacionals, i els camèfits, als deserts més secs.
Jordi Corbera, a partir de fonts diverses
Totes les plantes necessiten aigua per a sobreviure; per això es comprèn que, als deserts, les adaptacions vegetals s’orientin a obtenir el màxim benefici de la poca aigua disponible i a reduir al mínim les pèrdues d’aigua per evapo-transpiració. Hi ha plantes que, enfront de l’aridesa, adopten estratègies passives: eviten la sequera i resten més o menys inactives durant la calorosa estació seca. N’hi ha que adopten estratègies actives: resisteixen la sequera i es mantenen en activitat durant tot l’any. Els tipus biològics més freqüents en deserts i subdeserts (teròfits, gran part dels geòfits, hemicriptòfits i alguns camèfits) corresponen a espècies d’estratègies passives; només camèfits i faneròfits han d’acudir a estratègies actives enfront de l’aridesa climàtica.
Les plantes són més vulnerables a la sequera mentre germinen i s’estableixen. Algunes, les anomenades efímeres, són capaces de completar el seu cicle vital en les poques setmanes que dura l’estació plujosa i, efectivament, de vegades el cicle vital ha de ser realment molt curt. Hi ha gramínies del gènere Boerhavia i zigofil·làcies del gènere Tribulus, per exemple, que triguen només entre 8 i 15 dies a germinar i donar fruit. Hi ha plantes herbàcies perennes que, sobretot en condicions d’estrès, es comporten com a efímeres o com a molt poden produir llavors un parell d’anys. En canvi, els arbres i els arbustos necessiten anys per a arribar a l’estadi reproductiu i, per tant, estan més exposats als riscos de la pastura i de la manca de pluges abans no assoleixen la maduresa i poden començar a reproduir-se. Fins i tot, una pluviositat apropiada no és suficient per a assegurar l’èxit, per exemple, quan es pretenen introduir espècies llenyoses econòmicament aprofitables en una nova àrea. A la zona del Sahel, al Sàhara meridional, per exemple, l’elevat dèficit de saturació i la baixa humitat relativa hi impedeixen cultivar —excepte a la zona costanera occidental, on les condicions són lleugerament més favorables— tota una sèrie de plantes que podrien ser molt útils, com per exemple el nopal (Opuntia), els salats (Atriplex) o les acàcies australianes anomenades “wattles” (Acacia del subgènere Phyllodinae).
Les formes de germinació i de dispersió
Els cicles de tipus anual són un mitjà per a assegurar el futur genètic d’una espècie, o sigui, la dispersió i la supervivència de les generacions futures a través de les llavors. Entre les estratègies que han desenvolupat les plantes del desert per assegurar la supervivència i dispersió de les llavors n’hi ha que inclouen ritmes endògens d’aquestes que controlen la proporció de llavors que poden germinar en cada moment de manera que els règims de temperatura i fotoperíode abans i durant la germinació siguin més favorables. Molt sovint les llavors es desprenen a principi o durant l’estació seca, bé que algunes resten a la planta fins que les condicions són adequades per a germinar.
Les poàcies o gramínies constitueixen una família nombrosa i amb gran èxit evolutiu que té representants a totes les regions climàtiques del món, sovint amb un paper destacat, quan no dominant, en el paisatge. En un ambient semiàrid, les poàcies anuals, com Sporobolus festivus al Sahel, són les primeres que apareixen en començar l’estació plujosa. De vida curta, S. festivus completa el seu cicle vital en poques setmanes, abans que puguin ofegar-ne el creixement les poàcies perennes que es desenvolupen més tard. Les plantes herbàcies anuals solen ser espècies oportunistes petites capaces d’ocupar un nínxol adequat mentre les condicions per a créixer siguin bones. Poden sobreviure a la llarga estació seca i als anys de sequera prolongada en forma de llavors, i amb l’arribada de les pluges germinen i completen el seu cicle vital abans que les condicions empitjorin. Aquest sistema de vida està molt ben adaptat als ambients desèrtics, tot i que sovint mostra poques adaptacions a la sequera.
La capacitat de produir llavors, amb requeriments de diversos graus d’humitat i altres tractaments abans de germinar, està molt desenvolupada entre les plantes del desert. En l’aïzoàcia Mesembryanthemum nodiflorum són les llavors terminals de la càpsula les primeres a ser alliberades; a mesura que la planta es va humitejant, les segueixen les llavors centrals, i finalment les de la part basal. Les llavors terminals germinen al cap d’uns quatre anys, mentre que les altres requereixen uns anys de maduració abans de germinar. Al Namib, les llavors de l’aïzoàcia Delosperma pergamentaceum germinen gairebé totes alhora. Aquest comportament implica un risc considerable, perquè podria ser que no plogués prou i les llavors no arribessin a germinar. De fet, però, es tracta d’una espècie suculenta molt longeva que creix lentament, i amb poques llavors ja en té prou per a mantenir la població. Els aquenis de la composta anual Asteriscus pygmaeus queden a la planta morta embolicats per unes dures bràctees involucrals. Al llarg d’uns anys, amb la pluja, es van alliberant centrípetament del capítol sec. Altres compostes, com per exemple diferents espècies de Crepis, Geropogon i Picris tenen aquenis de morfologies diferents (heterocàrpia), uns de centrals amb vil·là i uns altres de perifèrics amb un vil·là rudimentari. Els últims no es podran dispersar fins que arribi la ventosa estació plujosa, i no aniran a parar gaire lluny de la planta mare.
L’amficàrpia és la capacitat de produir inflorescències de dos tipus, una d’aèria normal i una altra de subterrània més reduïda. La primera està adaptada a la dispersió a gran distància, mentre que la segona es desenvolupa aproximadament al mateix lloc. Són amficàrpiques la poligonàcia Emex spinosa, una herba anual sàharo-aràbiga i mediterrània, i també en la asteràcia anual Gymnarrhena micrantha. Emex asperatus i E. cylindricus són dues herbes amficàrpiques anuals australianes que, juntament amb la pantropical E. desvauxii, produeixen clistògens ben protegits per una beina, i es disper-sen mitjançant les tiges, que es desarticulen al nivelldel nus inferior. Les fabàcies anuals Ononis sicula i Trigonella arabica, quan plou molt, produeixen llavors amb embolcalls perfectes, mentre que les llavors que maduren a la mateixa planta mare quan plou poc tenen proteccions menys permeables. Una cosa semblant succeeix amb moltes papilionàcies dels gèneres Astragalus, Medicago i Trigonella, que tenen fruits amb moltes llavors, però de les quals cada any només n’arriba a germinar una.
La presència d’inhibidors de la germinació, tant als propàguls com a la fullaraca, és una manera de retardar la germinació fins que ha plogut prou per alixiviar les substàncies al·lelopàtiques del sòl i, per tant, per a proveir humitat suficient. A Israel, l’asteràcia Pectis papposa necessita com a mínim 10 mm de pluja per a germinar. S’han trobat inhibidors de la germinació a les valves dels fruits de la zigofil·làcia Zygophyllum dumosum, a les bràctees dels salats (Atriplex), a les fulles del perigoni o embolcall floral de la poligonàcia Rumex roseus i a la coberta de les llavors de la fabàcia Colutea istria. L’àcid nordihidroguaiarètic, present a les fulles de la zigofil·làcia Larrea tridentata, possiblement és l’agent responsable de l’efecte al·lelopàtic que pateixen les plantes que volen establir-se al voltant d’aquesta espècie (incloses les mateixes plàntules), amb la qual cosa s’elimina qualsevol amenaça a la supervivència.
Les formes de resistència
Les plantes perennes que broten sota la superfície del sòl i que durant l’estació seca perden la part aèria, de manera que només poden sobreviure gràcies a l’aliment acumulat en els òrgans de reserva, són anomenades geòfits. El seu cicle vital els permet desenvolupar arrels, fulles i flors durant la curta estació de creixement, quan les condicions d’humitat i de temperatura són òptimes; durant la resta de l’any es limiten a sobreviure a la calor i a l’aridesa extremes. Si plou poc, aquest estadi de repòs pot allargar-se tot un any o fins i tot més. Però, curiosament, hi ha alguns geòfits que poden créixer i florir abans que comencin les pluges. N’hi ha, com les liliàcies dels gèneres Allium, Asphodelus, Bellevalia, Ornithogalum o Tulipa, que desenvolupen les fulles i les flors simultàniament. D’altres, com les també liliàcies Scilla o Urginea, desenvolupen les fullesi les flors en estacions diferents. Entre els òrgansde reserva perennes es troben bulbs tunicats (per exemple els de la liliàcia Hesperocallis undulata), bulbs compactes (per exemple els de les iridàcies dels gèneres Crocus, els safrans, i Gladiolus, els gladiols), rizomes (com els de la gramínia Ammophila arenaria) i tubercles (com els de la icacinàcia Pyrenacantha malvifolia).
Els bulbs tunicats són brots molt modificats que formen un òrgan subterrani, carnós i perenne, la massa del qual és constituïda per bràctees o per la part basal de fulles disposades al voltant d’una gemma apical central amb fulletes immadures, la futura flor, i unes arrels adventícies rudimentàries a la seva base. Es troben ben representats entre les famílies de les liliàcies, de les amaril·lidàcies i d’altres monocotiledònies emparentades.
Els bulbs compactes són òrgans subterranis perennes, curts i engruixits, que desenvolupen fulles i flors a partir d’una o més gemmes axil·lars; la planta començarà a créixer a expenses de les reserves alimentàries acumulades en el bulb. Les esquames protectores de color marró que envolten el bulb sòlid són restes de les fulles basals d’anteriors estacions. Es donen sovint en les iridàcies.
Els rizomes es troben tant entre les dicotiledònies com entre les monocotiledònies. Són tiges subterrànies horitzontals i normalment ramificades que treuen arrels al nivell dels nusos i actuen com a agents de propogació vegetativa. Els rizomes poden ser allargats en forma de corda, com en moltes herbàcies perennes, o bé gruixuts i carnosos, com per exemple en les tifàcies del gènere Typha, les bogues. La gramínia Sporobolus rigens, que creix a les dunes mòbils de la regió del “monte”, a l’Argentina, té rizomes de més de 15 m que se situen entre els 25 i els 35 cm superiors del sòl, mentre que les seves arrels poden fer 40 cm de llargada i arribar a 70 o 80 cm de profunditat. Les puntes dels rizomes presenten gemmes resistents que perforeni penetren al sòl. Els rizomes es poden estendre fins a 6 m en un any, i donen brots aeris cada 18 o 20 cm.La seva capacitat de rebrotar després de quedar enterrats per la sorra fan de S. rigens una de les plantes més útils per al control de les dunes mòbils.
També es donen tubercles en diferents espècies, tant de dicotiledònies com de monocotiledònies dels deserts i els subdeserts càlids. Els tubercles poden ser caulinars (derivats de la tija) o radicals (derivats de l’arrel); els primers es distingeixen dels segons per la presència de gemmes o ulls. Serien exemples típics de tubercles caulinars la icacinàcia Pyrenacantha malvifolia i la dioscoreàcia Dioscorea elephantipes, els tubercles comestibles i d’escorça gruixuda de les quals deriven del primer entrenús de la tija. Els tubercles radicals, sense gemmes, són simples òrgans de reserva on s’acumula midó, aigua o totes dues coses. El “mosmote” o “pochote” (Ceiba acuminata), del desert de Sonora, i C. parvifolia, de les regions semiàrides de Puebla i Oaxaca, a Mèxic, per exemple, són bombacàcies que presenten tubercles radicals amb midó; malgrat el seu escàs valor nutritiu, es produeixen en quantitats prou grans per a constituir un dels aliments bàsics de les poblacions locals.
Finalment, es pot fer esment de la peculiar forma de creixement d’algunes espècies del gènere Eucalyptus dels subdeserts australians, a les plàntules de les quals apareixen petites protuberàncies a l’angle que queda entre les dues primeres fulles (cotilèdons). Aquestes petites protuberàncies creixen cap avall fins a embolicar la tija (hipocòtil) i acaben formant una mena de rabassa o tubercle llenyós a partir del qual es desenvoluparan les branques aèries, que s’obriran com una ombrel·la per formar una capçada prima, còncava o aplanada. Algunes formes de creixement semblants, bé que amb brots de vida curta o que s’assequen anualment, han evolucionat independentment en una trentena de famílies de plantes de l’Àfrica tropical i subtropical, segurament com a resposta a condicions edàfiques poc favorables, sense que el foc en sigui la causa principal.
Les estratègies d’optimització dels recursos hídrics
Les precipitacions escasses, les temperatures altes i l’elevat potencial d’evapo-transpiració que hi va associat obliguen en general les plantes del desert a economitzar l’aigua per tal de sobreviure. L’optimització en l’ús dels recursos afecta tant les formes de captació de l’aigua com els sistemes anatòmics i fisiològics per a treure’n el màxim rendiment.
Els sistemes de captació d’aigua edàfica i freàtica
Els freatòfits, arbres i arbustos amb arrels que arriben al nivell freàtic, com el “tamarugo” (Prosopis tamarugo), exploten les reserves d’aigua del subsòl. Mentre s’excavava el canal de Suez es van trobar arrels d’“afadar” (Acacia tortilis) a 30 m de profunditat, i se sap que l’arrel principal d’algunes espècies americanes de Prosopis pot arribar als 80 m. A la regió de Marràqueix, al Marroc, excavant un pou, s’han trobat a 60 m de fondària arrels d’1 cm de gruix de la ramnàcia Ziziphus lotus.
Si bé hi ha freatòfits que es converteixen en arbres i arbustos alts, altres plantes llenyoses de les regions àrides tenen òrgans subterranis massissos, però els brots que afloren a la superfície són anuals o de vida molt curta. És el cas de la mimosàcia arbustiva Andira laurifolia varietat laurifolia de la “caatinga” nord-oriental del Brasil. Aquesta espècie fa uns brots amb fulles no gaire llargs, d’uns 30 cm d’alçària, que li permeten reduir la transpiració al mínim, i desenvolupa òrgans caulinars de reserva en els 2 m superiors del sòl, mentre les arrels absorbents baixen uns 18 m fins al nivell freàtic. També és espectacular el cas del “kad” (Acacia [=Faidherbia] albida), que explota tots els aqüífers que es troben fins a 80 m de profunditat. A les plàntules dels freatòfits els cal desenvolupar arrels principals profundes tan ràpidament com els sigui possible, ja que si no poden arribar a les capes humides del subsòl abans de l’estació seca, no sobreviuran. De fet, molt poques plàntules arriben a la maduresa, fins i tot en anys favorables. En alguns deserts, com el Sàhara i el desert d’Aràbia, l’aigua subterrània es troba a tanta profunditat que les arrels no hi poden arribar; en altres, com al desert d’Atacama, que es va originar per un sollevament del llit de la mar, l’elevada salinitat no permet a les plantes de sobreviure, tot i la presència d’un nivell freàtic elevat alimentat per la neu que es fon dels Andes.
Adaptacions de les plantes
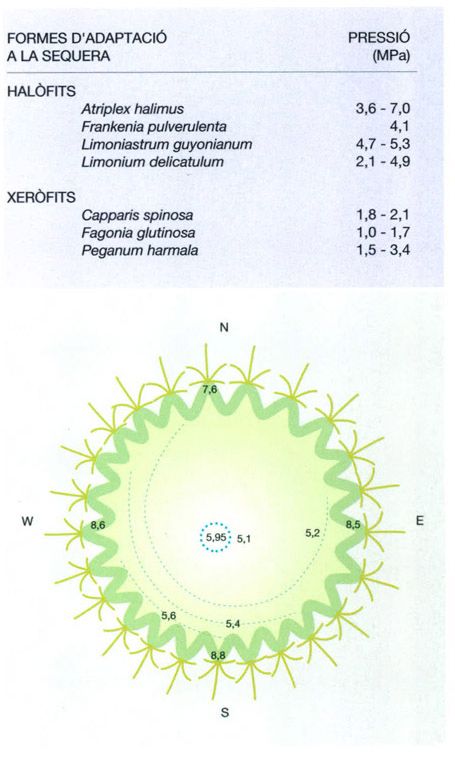
Jordi Corbera, a partir de Walter / Breckle, 1984 i dades proporcionades per l’autor
L’elevada pressió osmòtica dels teixits de les arrels i els brots de les plantes xeròfiles és una adaptació fisiològica que permet millorar l’absorció d’aigua per les arrels, i està inversament relacionada amb la disponibiltat d’aquest element. Així mateix, les plantes que viuen en medis salins (halòfits) tenen una pressió osmòtica més elevada que els xeròfits d’hàbitats no salins, cosa que s’observa en comparar alguns valors presos al desert del Sàhara (a baix). D’altra banda, la pressió osmòtica no és pas la mateixa a totes les estructures vegetals, tal com mostren uns estudis sobre les cactàcies de l’Amèrica del Nord. Els teixits corticals dels cactus presenten valors més alts que els teixits assimiladors interiors, mentre els mínims es donen al parènquima de reserva. Els mateixos estudis demostren que la pressió osmòtica també varia a les diferents parts de la planta segons cap on estiguin orientades. Els valors més alts es troben a la banda que mira al SW, ja que en dies assolellats les temperatures màximes i el dèficit de saturació de l’aire es donen a partir de migdia, i a l’hemisferi nord és la cara SW la que queda exposada al sol de la tarda. També hi ha una clara relació entre l’estructura i la pressió osmòtica, tal com s’observa en representar les línies isosmòtiques (valors en atmosferes) sobre una secció de Ferocactus wislizenii (a dalt): quan més elevats són els valors de la pressió osmòtica, més juntes es troben les costelles i més ben desenvolupat està el sistema vascular.
Però no totes les plantes del desert són freatòfites. Les estratègies alternatives per a les plantes perennes no-freatòfites dels deserts i els subdeserts consisteixen, bé en un sistema radical extens, just per sota la superfície, que els permet aprofitar qualsevol petit ruixat, o bé en una combinació d’arrels poc profundes i una arrel principal moderadament llarga amb ramificacions laterals, la qual els serveix per a absorbir l’aigua de les precipitacions fortes que s’escola avall pel perfil del sòl. La primera d’aquestes estratègies és típica de moltes plantes anuals i efímeres; la segona, de molts arbres, arbustos i herbes perennes, tot i que diverses espècies arbustives suculentes, principalment de la família de les cactàcies, tenen un sistema radical poc profund. Per exemple, el “saguaro” (Carnegiea gigantea) és un cactus columniforme que pot arribar a fer més de 12 m d’alçària, tot i que les seves arrels sovint no arriben a més d’1 m de profunditat. De tota manera, desenvolupa nombroses arrels laterals poc profundes que s’estenen més de 30 m i li permeten captar l’aigua de la pluja més insignificant en una superfície força extensa.
Les plantes que han pogut arrelar, independentment de si es tracta d’arbres, arbustos o herbes, podran sobreviure si l’àrea de captació d’aigua que cobreix el seu sistema radical els proporciona humitat suficient per a créixer. Això determina la densitat de la vegetació, de manera que un sistema radical que s’estengui per un radi de 30 m necessitarà un espai com a mínim de 60 m entre els arbres i els arbustos amb un sistema radical de dimensions similars. Això no vol dir que plantes amb sistemes radicals diferents o amb diverses necessitats estacionals d’humitat no puguin créixer en aquesta àrea de captació. El sistema d’arrels laterals pot ser efectiu en sòls nus, com passa, per exemple, als bosquetons de “mulga” (Acacia aneura) australians. La presència de plantes perennes, anuals i efímeres, tant si hi ha una matriu arbòria o arbustiva com si no, és un reflex de la quantitat i la distribució de les precipitacions, com també de la competència de les arrels en l’espai i en el temps.
Gràcies al seu port, moltes plantes llenyoses poden obtenir humitat addicional concentrant l’aigua de l’atmosfera a la capçada. L’aigua de les pluges o de les rosades intenses que es condensa a la capçada es pot canalitzar cap al sòl a través de les fulles, les branques i la tija. Un cop ha penetrat al sòl, és absorbida per les arrels i canalitzada altra vegada cap a la capçada. Les poàcies i altres plantes herbàcies poden créixer de manera relativament esponerosa al microambient creat a la base d’arbres i arbustos, en part per efecte de l’ombra, que crea un ambient una mica més frescal, i en part gràcies a un règim d’humitat més alt a resultes del flux caulinar abans que sigui absorbit per les arrels de les plantes hostes. Aquest exemple il·lustra, també, com les herbes amb un sistema radical poc profund poden competir per l’aigua amb els arbres i els arbustos d’arrels més profundes.
Les plantes que resisteixen el vent es converteixen, per aquesta mateixa raó, en punts on s’acumula la sorra. Conseqüentment, moltes plantes han d’adaptar el creixement al nivell canviant del sòl si no volen quedar enterrades. Així, per exemple, en els casos de la llemenosa (Artemisia campestris), de l’asteràcia Rhanterium suaveolens o del siscall (Salsola vermiculata), després d’una moderada acumulació de sorra, la planta mare acaba separada en diverses plantes funcionalment independents. Altres espècies, com el “markh” (Leptadenia pyrotechnica), de la família de les asclepiadàcies, desenvolupen arrels adventícies a les tiges que han quedat colgades per tal d’adaptar-se a la humitat i a la disposició canviant de les dunes. Rhanterium suaveolens pot fer una cosa semblant, i les seves arrels adventícies acaben actuant a la llarga com a arrels principals que formaran plantes funcionalment independents.
Semblantment, les arrels adventícies de la base dels estolons destinats a quedar colgats permeten a la compacta i rizomatosa poàcia perenne Stipagrostis pungens superar qualsevol acumulació de sorra. Allà on aquesta espècie és dominant, després que hagi desaparegut Rhanterium, es formen muntells de sorra fins de 2 m d’alçada. Les arrels de S. pungens queden recobertes en tota la seva extensió per una capa de sorra. La capa pilífera de l’arrel actua com una glàndula secretora de mucilag, que ajuda la seva punta a obrir-se camí entre la sorra; alhora, els grans de sorra, sucats pel mucilag, s’adhereixen entre ells i amb l’arrel fins a formar una beina de sorra. Les cèl·lules de la capa pilífera, aleshores, es transformen en pèls absorbents que no secreten mucilag, però que serveixen com a òrgans d’absorció. La beina sorrenca no tan sols protegeix l’arrel i els pèls radicals de la calor i la sequera, sinó que també augmenta la capacitat d’absorció de l’arrel. És més, un estudi recent dut a terme als Estats Units ha detectat la presència de nitrogenasa associada amb la beina sorrenca de determinades poàcies americanes, entre elles Aristida purpurea, Oryzopsis hymenoides i Stipa comata, bé que encara no s’ha demostrat que aquestes espècies puguin fixar nitrogen. Se sap que les beines sorrenques apareixen també en moltes altres gramínies del desert, com per exemple l’espart bord o espart albardí (Lygeum spartum), Panicum turgidum o Stipagrostis [=Aristida] obtusa. A les herbàcies anuals Bromus tectorum, el margall bord (Hordeum murinum) i Schismus barbatus [= S. calycinus], però, la coberta de sorra queda retinguda pels pèls absorbents i no pas per una secreció mucilaginosa. Si en altres famílies també es forma una beina sorrenca és una cosa que encara està per investigar; però al Sahel s’ha observat, sense cap mena de dubte, en papilionàcies del gènere Tephrosia.
La configuració foliar i l’estalvi d’aigua
Una adaptació molt lògica per a reduir les pèrdues d’aigua és la renovació dels òrgans responsables de la major part d’aquestes pèrdues, és a dir, de les fulles. Les fulles normalment cauen quan l’aigua que absorbeixen les arrels no és suficient per a reemplaçar la que es perd per evapo-transpiració durant el dia. Això sol ocórrer a principi de l’estació seca, però es pot donar també en períodes de sequera prolongada. Tot i que la caiguda anual de les fulles al final de l’època de creixement és un fenomen típic de les espècies caducifòlies, les espècies perennifòlies, que en condicions normals conserven les fulles durant tot l’any, poden perdre també les fulles sota condicions de fort estrès hídric. En absència de les fulles, una part del còrtex pot encarregar-se de la fotosíntesi, com passa per exemple en les quenopodiàcies Haloxylon salicornicum i Anabasis articulata.
L’aïllament fisiològic i la mort d’una branca o d’un grup de branques carregades de fulles és un mètode encara més dràstic per a reduir les pèrdues d’aigua. És el que fa la quenopodiàcia Atriplex torreyi, un arbust perennifoli que té la tija estriada per efecte de la separació dels radis corticals i d’una activitat desigual del càmbium. Aquesta peculiar estructura de la tija li permet deixar fisiològicament aïllades algunes branques, i concentrar l’aigua només en una sola o en poques. De manera semblant, al Nègueb, els troncs de la botja pudent (Artemisiaherba-alba) tenen unes bandes de cèl·lules suberitzades que divideixen la planta en unitats, cadascuna amb el seu propi sistema radical. Aquelles unitats que durant un any especialment sec no puguin proporcionar aigua suficient, moriran. La mateixa adaptació es troba en la composta Achillea fragrantissima i en l’harmalà (Peganum harmala).
Les pèrdues per evapo-transpiració a través de les fulles també es poden limitar amb un engruiximent de la cutícula, que sovint va acompanyat d’un engruiximent de les parets de les cèl·lules epidèrmiques i fins i tot de les subepidèrmiques. La secreció de cera per la superfície externa de la fulla es pot produir en quantitats prou importants perquè tingui un interès comercial, com passa amb la palmera Copernicia prunifera.
També la disminució de la superfície foliar pot reduir la pèrdua d’aigua. A les plantes microfil·les aquesta reducció pot ser extrema, com a les èfedres (Ephedra) i en diverses quenopodiàcies halòfiles com les cirialeres (Arthrocnemum, Salicornia) i les barrelles (Salsola) en què les fulles s’han reduït a estructures escamoses i la fotosíntesi ha estat assumida per la tija. No resulta tan dràstic el dimorfisme foliar, que consisteix en el fet que les fulles de l’estació seca són considerablement més petites que les de l’estació humida. En les acàcies australianes del subgènere Phyllodinae els individus adults ja no treuen fulles i la funció clorofíl·lica la fan els fil·lodis, que normalment consisteixen en uns pecíols foliars plans i elàstics.
La pubescència i la pilositat redueixen considerablement la velocitat del vent sobre la superfície de les fulles i, conseqüentment, també les pèrdues per evapo-transpiració. Tanmateix, en alguns casos, en eliminar els pèls, la transpiració disminueix. Per exemple, els tricomes de Crassula absorbeixen fàcilment les gotetes d’aigua superficials, de manera que no és tan sorprenent que, de vegades, en eliminar els pèls, es redueixi la superfície de transpiració. Els pèls, però, també tenen altres funcions, com dispersar la radiació reflectida pel sòl, de manera que refreden la fulla; també protegeixen la planta dels insectes i altres animals. Les fulles i les tiges de moltes quenopodiàcies halòfiles són recobertes de pèls vesiculars que en les fulles joves verdes són plens d’una solució salina. Durant l’estació seca els pèls vesiculars s’assequen, les sals que tenen a l’interior precipiten i produeixen un efecte reflector, de manera que disminueix la quantitat de radiació solar que arriba a la planta.
Finalment, els estomes que se situen al revers de la superfície foliar queden menys exposats als efec-tes dels vents, de manera que les pèrdues per evapo-transpiració no són tan grans. Les fulles cargolades o doblegades aconsegueixen uns resultats similars en protegir els estomes. Moltes herbàcies del desert, per exemple, no tan sols cargolen les fulles sota condicions d’estrès hídric, sinó que tenen estomes només a l’anvers de la fulla i encara enfonsats.
El sentit hídric de l’espinescència i de la suculència
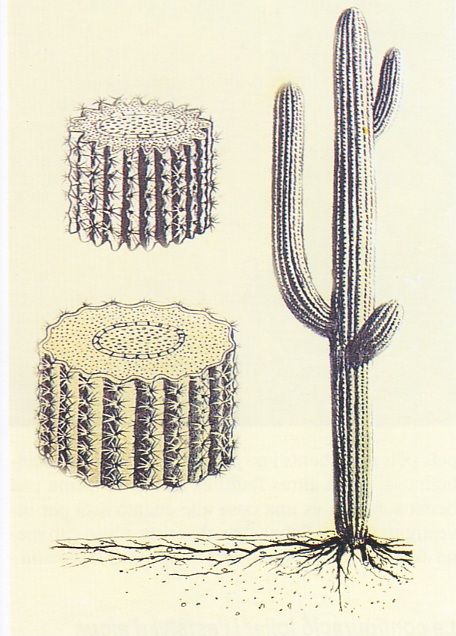
Moltes espècies de cactus presenten una disposició acanalada en acordió, ja que això els permet inflar-se i desinflar-se segons la quantitat d’aigua que contenen, tal com il·lustra aquest cartell informatiu sobre el cactus “saguaro” (Carnegiea gigantea) del desert de Sonora, a Arizona (EUA). El “saguaro” es va botint a mesura que absorbeix aigua. Els seus plecs en acordió li permeten expandir-se al voltant d’un esquelet llenyós, massís però elàstic, constituït per una trama cilíndrica formada per grups de feixos vasculars que s’anastomosen fins a mitja alçada i a partir d’allà queden lliures. La gran capacitat d’emmagatzemar aigua i el creixement lent d’aquest cactus li permeten donar fruit cada any, més o menys independentment de la sequera. El contingut d’aigua dels seus teixits representa entre el 75 i el 90% del pes fresc.
Ramon Folch / ERF
Les espines foliars són fulles modificades (aquest seria el cas de les espines de les cactàcies) o bé parts d’una fulla (per exemple, estípules en els casos de les mimosàcies dels gèneres Acacia i Prosopis). Aquestes modificacions redueixen la superfície foliar i per tant les pèrdues per transpiració. En els cactus, la punta de les espines actua com un nucli on es condensen les gotetes d’aigua, que regalimaran fins a terra, i aleshores les arrels engruixides abans esmentades aprofiten aquesta humitat. Les formes juvenils del “saguaro” (Carnegiea gigantea) van armades amb espines punxegudes força llargues orientades cap avall que els protegeixen de la depredació dels animals petits; a mesura que la planta va creixent, les espines es van posant dretes i s’orienten cap enfora. Les espines i els agullons són una bona defensa contra els herbívors, tot i que en les plàntules joves solen mancar o bé són toves. Paradoxalment, les plàntules d’Acàcia i Prosopis, per exemple, són vulnerables als herbívors durant l’etapa de creixement, quan encara són susceptibles a la manca d’aigua i requereixen optimitzar la fotosíntesi d’un fullatge escàs mentre es concentren en el desenvolupament de les arrels.
La suculència és una adaptació de moltes plantes xeromòrfiques que permet emmagatzemar l’aigua al mateix temps que en redueix les pèrdues per evapo-transpiració. Els teixits de les tiges i de les fulles engruixides són formats en gran part per un parènquima de reserva translúcid que acumula aigua. La pèrdua d’aigua es redueix gràcies a la menor proporció superfície/volum que fa que quedi exposada una àrea més petita de l’òrgan. Les plantes amb aspecte de còdols vivents del Namib, com les aïzoàcies del gènere Lithops i altres com les fenestràries (Fenestraria), encara redueixen més la pèrdua d’aigua deixant que les seves tiges engruixides s’incrustin al sòl, de manera que només exposen les finestres de llum de les fulles.
Els òrgans en forma de tiges engruixides especialitzats en l’emmagatzematge d’aigua són corrents en agavàcies, aloeàcies, asclepiadàcies, cactàcies i crassulàcies, en algunes aïzoàcies (Lithops, Nananthus, etc.), apocinàcies (Adenium), euforbiàcies, etc., com també en quenopodiàcies halòfiles dels deserts salats. La suculència s’ha desenvolupat paral·lelament en una sèrie de famílies no emparentades, les semblances externes de les quals no estan necessàriament relacionades amb les estructures internes. Per exemple, les cactàcies columniformes del Nou Món s’assemblen a les euforbiàcies columniformes (Euphorbia) del Vell Món; les fouquieriàcies americanes s’assemblen a les didieràcies de Madagascar, i les agavàcies (Agave) del Nou Món s’assemblen a les aloeàcies (Aloe) del Vell.
En els sòls salins, especialment a les regions àrides i semiàrides, on l’evaporació és superior a la precipitació, les plantes no sols han de lluitar contra l’aridesa, sinó també contra la salinitat. El mecanisme que fan servir per a regular l’absorció de sals encara no es coneix del tot. En la major part dels casos implica unes adaptacions osmòtiques molt desenvolupades que permeten mantenir la tensió necessària per a l’absorció d’aigua. Les plantes halòfiles, que són les que creixen en sòls d’aquesta mena, acumulen grans quantitats de ions sodi i clor que els permeten mantenir uns potencials interns baixos i evitar un dèficit d’aigua. La quantitat de sal que es transporta fins als brots és superior a la que es requereix per a regular la turgència. Les glàndules de la sal i els pèls esquamosos de les fulles de moltes halòfiles segreguen activament els ions sodi i clor, amb la qual cosa eliminen l’excés de sals. És el que fan, per exemple, les poàcies dels gèneres Aeluropus i Distichlis, les franqueniàcies del gènere Frankenia, les primulàcies del gènere Glaux, les plumbaginàcies dels gèneres Limonium i Limoniastrum, les tamaricàcies del gènere Tamarix i diferents espècies de la família de les quenopodiàcies, en moltes de les quals, com per exemple les dels gèneres Arthrocnemum, Atriplex, Cornulaca, Haloxylon, Salicornia, Suaeda, etc., la suculència sembla també un mitjà per a reduir la concentració de sals amb grans volums d’aigua.
Adaptacions de les plantes de deserts i subdeserts càlids

Jordi Corbera
La suculència, l’espinescència i la manca o reducció extrema del sistema foliar caracteritzen els cactus i les plantes cactiformes, vegetals típics dels deserts i subdeserts càlids. Els cactus pròpiament dits pertanyen a la família de les cactàcies i són de procedència americana; llurs tiges suculentes i normalment fotosintetitzadores presenten canals i costes longitudinals, al llarg de les quals es disposen les anomenades arèoles, zones corresponents a les axil·les foliars, d’on, en el cas dels cactus, neixen les espines. Tot i compartir una mateixa conformació suculenta, espinescent i bàsicament afilla, els cactus (plana de l’esquerra) presenten morfologies i dimensions diversíssimes: n’hi ha de globulosos, com els Echinocactus de les zones desèrtiques nord-americanes i del N de Sud-amèrica, de la mida d’una pilota de bàsquet (se n’ha representat una arèola amb les espines); n’hi ha de pulvinulars, com les nombroses espècies de Copiapoa del desert d’Atacama (se n’ha representat una flor en secció i l’arrel napiforme, tan habitual en el cactus); i també n’hi ha d’arborescents monopòdics, com l’espectacular “candelabro” (Browningia candelaris) dels raiguers andins del desert de Tarapacá (Atacama), de fins a 5 m d’alt, d’arborecents simpòdics, com l’“organ pipe” (Lemaireocereus thurberi) de Sonora, poc espinós i de 2-3 m d’alt, o de columnars, com la pasacana d’Atacama (Trichocereus atacamensis), fortament espinós i pilós, de fins a 6-7 m d’alt. Una morfologia comparable és també adoptada per altres plantes desertícoles de les famílies de les euforbiàcies, aizoàcies, asclepiadàcies, agavàcies, paquipodiàcies, etc. (plana de la dreta), com és el cas, respectivament, d’algunes Euphorbia africanes arborescents, dels petits Lithops sud-africans de la mida i la forma d’un còdol, de les digitades Stapelia africanes, de les atzavares (Agave) nord-americanes d’espectaculars inflorescències, o dels estrambòtics Pachypodium malgaixos (se n’ha representat una fulla amb les estipules transformades en una característica espina bifida), que poden superar els 2 m.
La diversitat de la vegetació dels deserts i els subdeserts càlids
El que caracteritza per damunt de tot la vegetació dels deserts és la seva discontinuïtat en l’espai i en el temps. La vegetació de les zones desèrtiques es limita als punts baixos de la topografia i al llarg de la xarxa hidrogràfica; els relleus que separen les valls resten nus, fora d’una vegetació anual efímera, fugaç, a conseqüència dels escassos i breus episodis plujosos. Els subdeserts, en canvi, tenen una vegetació perenne més o menys difusa, poc o molt regularment repartida en el paisatge d’acord amb gradients geogràfics. La fisiognomia i la natura d’aquesta vegetació varien considerablement segons els climes, els sòls, la pressió antròpica i la gestió que se’n fa.
Els elements perennes de la vegetació dels deserts
La vegetació perenne pot variar als deserts segons la importància i la freqüència de l’escolament, tant si és autòcton o local com al·lòcton o procedent de pujols o muntanyes més o menys allunyades, i segons les crescudes. Pot ser constituïda per mates i arbustos, amb arbres o sense. Quan no hi ha arbres, pot prendre l’aspecte d’un erm com l’‘estepa’ amb artemísia (Artemisia) de l’Àfrica septentrional i del Pròxim Orient, l’‘estepa’ amb quenopodiàcies de l’Àfrica septentrional i del Pròxim Orient, l’‘estepa’ amb Rhanterium de l’Àfrica septentrional i del Pròxim Orient, el “karoo” amb aïzoàcies de l’Àfrica austral, etc; també pot adoptar l’aspecte d’una timoneda o erm arbustiu (Àfrica septentrional, Pròxim Orient, Sind, Thar, Àfrica austral, deserts americans, Austràlia) o, encara, el d’‘estepes’ o sabanes arbrades (Àfrica septentrional, Pròxim Orient, Sahel, Àfrica oriental i deserts americans). Quan n’hi ha, els arbres pertanyen generalment al gènere Acacia (A. ehrenbergiana, A. tortilis raddiana, al Sàhara i als deserts d’orient; A. gerrardii, al Sinaí i al Níger; A. nilotica, al Sind; A. karroo, A. giraffae, A. hematoxylon, A. erioloba, A. mellifera subspècie detinens, A. reficiens, A. [=Faidherbia] albida, i també Dichrostachys cinerea, a l’Àfrica austral; Acacia tortilis, A. nilotica, A. bussei , A. mellifera, A. asak, A. nubica, A. reficiens, a l’Àfrica oriental).
Els deserts sud-africans posseeixen, per altra banda, alguns arbres singulars, especialment àloes arborescents (Aloe arborescens, A. dichotoma, A. pillansii) i els paquipodis (Pachypodium succulentum, P. namaquanum); aquest últim presenta una curiosa convergència de forma amb el no menys curiós “bojum” (Fouquieria [= Idria] columnaris) del desert de Vizcaíno, a la Baixa California. S’observen també nombroses convergències de formes entre les eufòrbies cactiformes africanes i alguns cactus americans. El Namib se singularitza també per nombroses espècies suculentes, principalment aïzoàcies de port i flors molt ornamentals però també per tot un seguit d’altres espècies encara més sorprenents com la welwítsquia (Welwitschia mirabilis), una estranya coní-fera de fulles de creixement indefinit que és un veritable fòssil vivent vegetal; o l’arbust Myrothamnus flabellifolius, únic representant africà del seu gènere i de la família de les mirotamnàcies (que només comprèn una altra espècie, aquesta de les regions àrides de Madagascar), capaç de practicar una mena d’estivació que consisteix a assecar-se fins a semblar mort i revivificar-se així que plou; els arbres paquicaules que acumulen aigua al tronc com el “moringa” (Moringa ovalifolia), de la família de les moringàcies o la burseràcia Commiphora saxicola; la crassulàcia arborescent Cotyledon paniculata, les eufòrbies cactiformes Euphorbia virosa, E. damarana, E. gariepina i encara d’altres; les vitàcies Cyphostemma uter i C. currorii; la passifloràcia Adenia pechuelii; l’esterculiàcia Sterculia quinqueloba, o la portulacàcia Ceraria namaquensis.
Els deserts càlids nord-americans se singularitzen al seu torn per les iuques arborescents, conegudes també als Estats Units com a “Joshua trees” (Yucca schidigera, Y. valida, Y. brevifolia, Y. schottii), els cactus gegants columnars (Pachycereus pringlei, P. pecten-aboriginum, Lemaireocereus thurberi, Carnegiea gigantea, que és el “saguaro”), els diversos arbrissons amb aspecte arbori o “palos” (Cercidium floridum o “palo verde”, C. microphyllum o “palo azul”, C. sonorae > o “palo santo”, Olneya tesota o “palo hierro”), el “bojum” (Fouquieria [=Idria] columnaris ), espècies paquicaules com les anacardiàcies del gènere Pachycormus, o les burseràcies del gènere Bursera, acàcies com l’“uña de gato” o “catclaw” (Acacia greggii), el “palo blanco” (A. willardiana), l’aromer o “huizache” (A. farnesiana), el “Jerusalem thorn” o “mataburro” (Parkinsonia aculeata) i els prosopis, mesquites o “algarrobos” (Prosopis glandulosa,P. laevigata, P. torreyana, P. velutina).
La vegetació dels subdeserts
La vegetació dels subdeserts és força diferent en els marges temperats (mediterranis) dels deserts i en els seus marges tropicals. Els subdeserts de tipus mediterrani són constituïts principalment per formacions estepàries com els erms i les timonedes. Aquestes ‘estepes’ poden ser primàries, és a dir naturals, o secundàries, és a dir resultat de processos de degradació en formacions forestals: incendis, focs pastorals, desboscament, sobrepasturatge, artigament. Una gran part dels erms de l’Àfrica septentrional i dels erms i timonedes de la Península Ibèrica, fins a l’entorn de la isohieta de 200 mm anuals de pluja, són de natura secundària, originades a partir de boscos de pi blanc (Pinus halepensis), de savines (Juniperus phoenicea, J. thurifera) o de tuia articulada (Tetraclinis articulata). Al Pròxim Orient, les ‘estepes’ situades per damunt de la isohieta de 200 mm, resulten de boscos oberts amb pi blanc o festuc de l’Atles (Pistacia atlantica), amb diferents espècies de festuc (Pistacia), ametllers (Prunus dulcis), i localment, roures o alzines (Quercus).
La vegetació dels subdeserts intertropicals és, generalment, una sabana herbosa amb arbres dispersos, arbustos i plantes herbàcies anuals o perennes. Les mates són rares o absents. Les sabanes sahelianes, per exemple, es componen de petits arbres espinosos espaiats (Acacia, Commiphora, Balanites, Ziziphus, etc.) i d’una catifa més o menys contínua on són predominants les gramínies anuals. Les sabanes de les zones àrides de l’Àfrica oriental i meridional i de l’Índia presenten la mateixa estructura però l’estrat herbaci és format de gramínies perennes, i no anuals, contràriament a l’indicat pel Sahel. Aquest fet és degut en part a la distribució de les pluges: concentrades en els tres mesos d’estiu al Sahel, repartides de quatre a vuit mesos a l’Àfrica oriental i a Kalahari; amb dues estacions plujoses en el primer cas i una sola en el segon.
Els subdeserts càlids del Nou Món es caracteritzen, com els deserts, per la presència i la dominància de grups taxonòmics importants propis de la flora neotropical com els mesquites o “algarrobos” (Prosopis), les cactàcies, les atzavares, les iuques, i un cert nombre d’altres tals com les “gobernadoras” o “creosote bushes” (Larrea), que particularitzen els paisatges. A l’Amèrica del Sud, aquest espai és ocupat pel “monte”, la “caatinga” i el “sertão”.
Les zones àrides australianes es caracteritzen per la dominància d’arbres i d’arbustos dels gèneres Acacia, Cassia, Eucalyptus i Melaleuca. Entre els més importants es troben la “mulga” (Acacia aneura) i grosses mates de gramínies perennes vulnerants, les “spinifex” (Triodia). Les zones àrides subtropicals i mediterrànies d’Austràlia es distingeixen de les sabanes tropicals per la dominància de nombroses quenopodiàcies arbustives halòfiles (Atriplex, Maireana) que constitueixen pastures apreciades pels bens sobre els terrenys més o menys salats.
La florida esporàdica: el cas del desert d’Atacama
La regió desèrtica peruano-xilena es caracteritza per un particular dinamisme ecosistèmic associat a les pertorbacions recurrents i irregulars en els corrents marins i atmosfèrics conegudes com el fenomen d’El Niño. Aquestes pertorbacions, quan es produeixen, faciliten la incursió cap a l’interior del continent de masses d’aire humit provinents de l’oceà Pacífic, amb el resultat que les terres àrides reben, en un breu període de temps, una quantitat de precipitació que pot superar en molt la seva pluviositat ordinària, fins i tot multiplicar-la per un factor significatiu. Amb les temperatures adequades (de 15° a 17°C), suficient humitat acumulada en sòls de textura arenosa i un banc de llavors, la regió desèrtica és capaç d’expressar durant l’estació de creixement (agost-novembre) el fenomen localment conegut com a “florecimiento del desierto”, l’espectacular florida gairebé simultània i de breu durada d’un gran nombre de plantes de flors vistoses. La regió sencera es cobreix d’un mantell bigarrat i dens de geòfits i d’anuals acompanyat del rebrot de l’estrat arbustiu. Dominen les espècies amb flors de tonalitats que van de blavoses a blanquinoses, de vermelloses a rosades, de fúcsies a liles, de grogues a ataronjades.
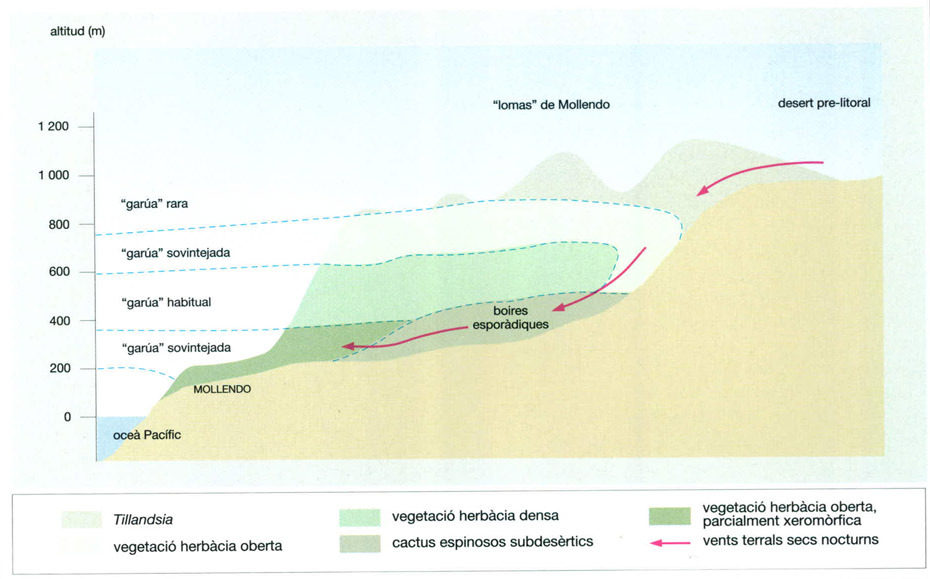
Un perfil esquemàtic de les “lomas” de Mollendo, al desert costaner peruà, mostra la freqüència de les boires (“garúa”) i la distribució de la vegetació al llarg d’un vessant que davalla de NE a SW des d’un altiplà desèrtic fins a la costa de l’oceà Pacífic. A la regió més elevada, on les boires són rares, la vegetació està dominada per bromeliàcies del gènere Tillandsia. Una mica més avall, les boires sovintegen més i apareix una vegetació herbàcia. Les espècies suculentes només creixen en biòtops concrets, allà on la zona de boires no queda totalment tancada, és a dir, a la vall on durant la nit bufen vents secs de les muntanyes que dispersen la boira. Les herbes tendres no poden resistir aquests vents que assequen l’ambient periòdicament. En canvi als cactus els van molt bé, ja que prevenen la infecció dels plançons per fongs i eliminen les herbes que els podrien fer la competència. Vora la costa, apareix una vegetació herbàcia oberta força xerofítica constituïda per espècies de boira.
Jordi Corbera, a partir de Walter / Breckle, 1984
Aquesta florida té lloc en la seva màxima expressió a les comunitats costaneres de “lomas”, on dominen herbes perennes i anuals de gèneres com Allionia, de la família de les nictaginàcies, Argylia, de les bignoniàcies, Caesalpinia, de les cesalpiniàcies, Calandrinia i Portulaca, de les portulacàcies, Cristaria, de les malvàcies, Eragrostis, de les poàcies, Leptoglossis, de les solanàcies, o Nolana, de les nolanàcies.
Més cap al S, amb precipitacions entre 80 mm (20°S) i 30 mm (27°S), la florida del desert s’expressa millor a les “lomas” costaneres i en els sòls arenosos dels “llanos” i “serranías” del desert interior transicional. Depenent de l’hàbitat (rocós o arenós) o del sector (costaner o plana interior), hi haurà un tipus d’associació dominant o un altre. Als “llanos” interiors (que cal no confondre amb les sabanes herboses d’altres regions tropicals més plujoses de l’Amèrica del Sud, també conegudes com a “llanos”), les més espectaculars per la seva combinació de colors corresponen a les associacions formades per la “malvilla” (Cristaria patens),malvàcia anual de flors rosades, la “pata de guanaco” (Calandrinia longiscapa), portulacàcia de flors de color fúcsia i la “flor del jote” (Argylia radiata), bignoniàcia de flors grogues. També cobreix una extensa zona de la plana central l’associació constituïda per la ja esmentada “pata de guanaco”, la Nolana paradoxa, que és una nolanàcia de flors blanquinoses, i Oenothera coquimbensis, una onagràcia de flors grogues, i l’associació formada per dues “malvillas” (Cristaria cyanea, de flors liloses, i C. patens, de flors d’un color fúcsia fort).
A la zona del litoral domina en alguns sectors el “quisco copiapoa” (Cristaria dealbata), amb una tija d’un estrany color gris platejat i flors grogues; en d’altres domina l’associació formada per “amancai” (la geraniàcia Balbisia penduncularis), “churqui” o “churco” (l’oxalidàcia Oxalis gigantea) i “alcaparra” (la cesalpiniàcia Cassia [=Senna] cumingii varietat coquimbensis), tots tres arbustos de flors grogues, acompanyats de cactàcies dels gèneres Eulychnia, Eriosyce, Neoporteria, Opuntia i Echinopsis. Arran de mar, amb sòls predominantment sorrencs, hi ha nombroses liliàcies bulboses, com les “añañucas” (Rhodophiala), de flors vermelles, grogues o blanques, els “huillis” o “cebollines” (Leucocoryne), de flors blanquinoses o blavoses, o els “azulillos” (Pasithea), de flors blaves; sovint en companyia dels “dondiegos de noche” (Oenothera), tan difosos a la jardineria dels països tropicals i mediterranis.
El desert florit adquireix la seva màxima expressió entre les latituds 27° i 29° S, on la diversitat florística és elevada. A l’herbassar abunden innombrables plantes anuals de variades famílies. Entre les liliàcies hi ha diverses “añañucas”, com l’“añañuca amarilla” (Rhodophiala bagnoldii) i l’“añañuca blanca” (R. laeta), juntament amb la seva congènere el “revienta ojos” (R. phycelloides); hi ha també la “mariposa de los molles” (Alstroemeria pelegrina) i els seus congèneres el “lirio del campo” (A magnifica), el “lirio amarillo” (A. kingii) i el “lirio rosado” (A. leporina), com també diferents “huillis” o “cebollines”, com el “huilli” de Coquimbo (Leucocoryne coquimbensis), L. narcissoides o L. purpurea i algun “azulillo”, com Pasithea coerulea.
A banda les liliàcies, hom troba aristoloquiàcies com l’“oreja de zorro” (Aristolochia chilensis), convolvulàcies com la “correhuela” (Convolvulus chilensis), nolanàcies com el “suspiro de campo” (Nolana paradoxa), malvàcies com les “malvillas” (Cristaria aspera, C. patens), violàcies com la “violeta del campo” (Viola asterias), aïzoàciescom l’herba gelada o “rocío” (Mesembryanthemumcrystallinum), asteràcies com el seneci Senecio brunonianus, papaveràcies com el “cardo blanco” (Argemone hunnemannii), solanàcies com la “mariposa blanca” (Schizanthus candidus), portulacàcies com la “pata de guanaco” (Calandrinia longiscapa), onagràcies com el “dondiego de noche” (Oenothera coquimbensis), franqueniàcies com la “hierba de salitre” (Frankenia chilensis), o euforbiàcies com el “lechero” (Euphorbia lactiflua).
Entre els arbustos es troben cesalpiniàcies com el “chañar” (Geoffroea decorticans), l’“alcaparra” (Cassia cumingii), el “retamo” (Caesalpinia angulicaulis) o l’“algarrobilla” (C. [=Balsamocarpon] brevifolium); mimosàcies com l’“espino” (Acacia caven); onagràcies com el “palo de yegua” (Fuchsia lycioides), geraniàcies com l’“amancai” (Balbisia peduncularis), boraginàcies com el “palo negro” (Heliotropium stenophyllum) o el “carbonillo” (Cordia decantra), compostes com la “flor del minero” (Centaurea chilensis), la “flor del minero rosada” (C. floccosa), la “coronilla de fraile” (Encelia canescens) o la “chamiza blanca” (Bahia ambrosioides), oxalidàcies com el “churco” (Oxalis gigantea), verbenàcies com la “verbena” (Junellia selaginoides), apocinàcies com el “cuerno de cabra” (Skytanthus acutus), nolanàcies com Nolana coelestis, i cactàcies dels gèneres anteriorment mencionats.
El desert florit és de curta durada. L’ecosistema inicia el retorn a la seva fase dominant a partir de mitjan octubre-novembre, quan la radiació solar, les temperatures i els vents del desert comencen a exercir la seva acció deshidratant. Al gener-febrer el paisatge ja ha tornat a la seva condició més característica, l’àrida.
La memòria d’un passat millor
Al Sàhara actual els arbres han quedat confinats gairebé exclusivament als oasis. Probablement, als oasis abans s’hi feien diferents espècies d’acàcia, i de caparidàcies dels gèneres Maerua i Capparis, juntament amb la palmera dum (Hyphaene thebaica), però la palmera datilera (Phoenix dactylifera) i altres plantes cultivades han reemplaçat els arbres originals. Una excepció notable a aquest confinament, l’havia constituït fins a la seva destrucció el solitari arbre del Ténéré, un exemplar d’“afadar” (Acacia tortilis subspècie raddiana), probablement resta d’un antic bosquetó. Havia pogut sobreviure, en un indret amb una pluviositat mitjana de menys de 10 mm, gràcies a la seva capacitat d’utilitzar l’aigua del nivell freàtic, que es troba a prop de 40m de profunditat. Considerat tabú pels caravaners tuaregs, protegit del brosteig i de la tala per a obtenir combustible, havia aconseguit sobreviure tot i que un vehicle militar li havia arrencat una part del tronc forcat. Malauradament, el 1973 va ser víctima d’un desafortunat accident d’un camió al qual ja no va poder sobreviure.
Altres relíquies històriques del clima, més humit que l’actual, que regnava al Sàhara al principi del Quaternari sobreviuen encara a les valls de les muntanyes saharianes. Als contraforts meridionals de les muntanyes de l’Atles, al Marroc, a la serralada de Mouydir, Tassili N’Ajjer i Ahaggar, a Algèria, i al mont Marra, al Sudan, hi creix una espècie d’olivera (Olea europaea laperrinei), que deu el nom al coronel Laperrine, el seu descobridor; en canvi, no apareix en altres àrees muntanyoses saharianes com el Tibesti, al Txad. Aquesta olivera formava part d’un conjunt d’espècies mediterrànies que penetraren al Sàhara central al principi del Quaternari, l’àrea de les quals s’ha anat reduint, a mesura que el clima s’ha anat tornant més i més àrid fins a quedar reduïda a les muntanyes esmentades. Si s’exceptuen les seves estretes fulles linears-lanceolades, la subspècie laperrinei s’assembla força a l’ullastre afro-asiàtic (O. europaea cuspidata), suposat parent de l’olivera cultivada (O. europaea europaea).
Una altra espècie relicta de la penetració plistocènica del Sàhara per la flora mediterrània és un xiprer endèmic, el xiprer del Tassili, xiprer de Duprez o “tarout” (Cupressus sempervirens dupreziana), quehavia estat descrit fins i tot com una espècie diferenciada independent (C. dupreziana). Es diferencia del xiprer comú (C. sempervirens sempervirens) pels seus característics brots aplanats i per la petitesa de les fulles. Els olis volàtils de les fulles de les varietats dupreziana i sempervirens són gairebé idèntics, cosa que confirma el seu estret parentiu taxonòmic. Els exemplars que encara viuen romanen confinats a unes quantes valls rocoses del Tassili N’Ajjer, a la veïna regió d’Ahaggar. Hi ha arbres nuosos i retorçats de més de 1 000 anys que poden arribar a 20 m d’alçària, i el seu tronc pot fer fins a 3 m de diàmetre. Antigament, la seva fusta era molt utilitzada pels tuaregs i, actualment, s’està intentant plantar aquesta espècie en altres regions, ja que és capaç de créixer en zones àrides i podria servir per a millorar la resistència a la sequera d’altres espècies emparentades.
A l’anomenada pampa del Tamarugal, al N del desert d’Atacama, a les depressions de gruixudes crostes salines (10-60 cm), hi creixen bosquets de “tamarugo” (Prosopis tamarugo). Tot i que el nivell freàtic es troba entre 1,5 i 40 m de profunditat, encara no és clar com aconsegueixen l’aigua aquests arbres. S’havia dit que durant la nit les fulles absorbien el vapor d’aigua a través dels estomes i n’excretaven l’excés per les arrels laterals, densament ramificades, aproximadament a 1 m sota la superfície del sòl. Aquest mecanisme, però, és fisiològicament impossible, i, per altra banda, les fulles no presenten cap adaptació per captar aigua. En realitat, el “tamarugo” és un freatòfit que absorbeix l’aigua a través de les arrels. La densa xarxa d’arrels laterals segurament correspon a una antiga zona absorbent que ha quedat per sobre del nivell de l’aigua freàtica; aquest nivell baixà quan els corrents d’aigua subterranis provinents dels Andes foren desviats cap a les ciutats mineres i costaneres de Xile. Tot i el seu comportament normal, el “tamarugo” no ha aconseguit gaire èxit fora del seu hàbitat original.