Característiques del grup
Els copèpodes constitueixen un dels grups animals que ha tingut més èxit evolutiu. No únicament és un grup molt diversificat (se’n coneixen prop de 8500 espècies), sinó que s’han sabut adaptar a pràcticament tots els ambients aquàtics. Entre les espècies de vida lliure, les espècies planctòniques constitueixen un dels grups d’organismes més abundants a la natura i, a més, tenen una gran importància a les cadenes tròfiques de tots els ecosistemes aquàtics, marins i continentals. Les espècies paràsites, bé que menys freqüents, també es troben molt diversificades (2500 espècies entre els pecilostomatoides i els sifonostomatoides) i, dins de la fauna marina, figuren entre els principals grups de vida paràsita.
Generalment són petits, de 0,5 a 2 mm de llargada, però n’hi ha alguns de més grossos, fins de 17 mm; entre els paràsits hi ha espècies d’una envergadura insòlita (fins a 32 cm), conseqüència de les ramificacions del cos que penetren dins dels teixits dels hostes. Els copèpodes tenen el cos dividit d’una manera peculiar dins dels crustacis, ja que únicament s’hi diferencien dues regions separades per una articulació. L’origen cefàlic, toràcic o abdominal dels metàmers que componen cada regió únicament es manifesta per l’estructura dels apèndixs respectius. A la part anterior del cos destaquen les antènules, mentre que la part posterior, mancada d’apèndixs, s’estreny progressivament i acaba en una furca. Els copèpodes no tenen closca ni ulls compostos. Des del punt de vista morfològic, però, cal distingir entre les espècies de vida lliure, que s’ajusten a aquesta descripció general, i les paràsites, que se n’allunyen pel fet d’haver sofert moltes modificacions secundàries, fins al punt d’haver-hi espècies que únicament es reconeixen com a copèpodes en els seus primers estadis larvaris. Per aquesta raó, tractarem separadament la morfologia i la biologia de les espècies lliures i les paràsites, i després farem l’anàlisi de les relacions que hi ha entre elles.
Morfologia
Espècies de vida lliure
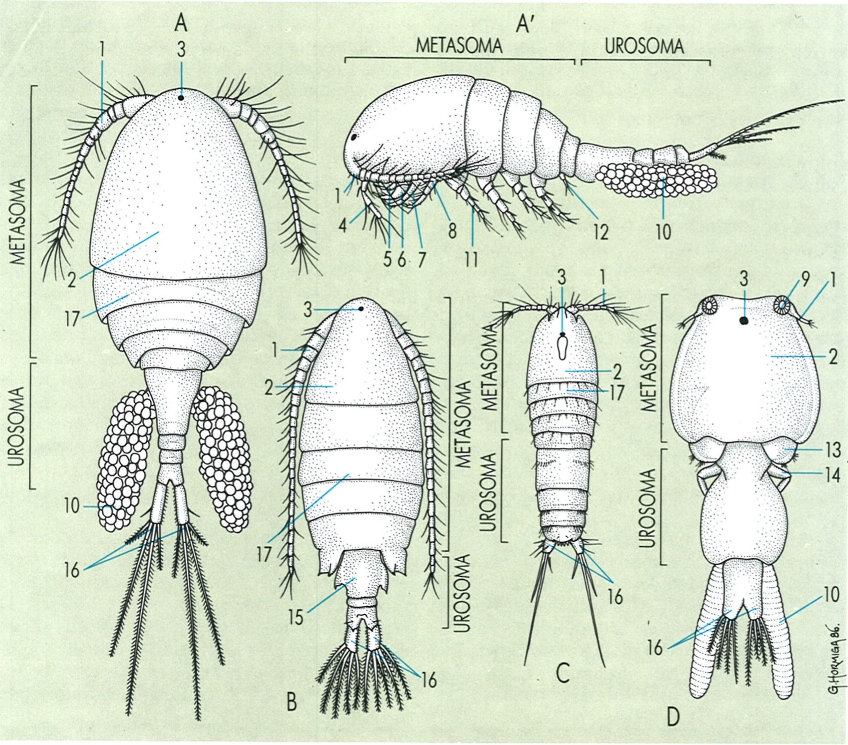
Aspecte general extern dels copèpodes, en un cas general, corresponent a un ciclopoide (× 100) en visió dorsal (A) i lateral (A’) i en els calanoides (B), harpacticoides (C) i sifonostomatoides (D) (× 70). Hom hi consigna: 1 antènula, 2 cefalosoma, 3 ull, 4 antena, 5 mandíbula, 6 maxíl·lula, 7 maxil·la, 8 maxil·lípede, 9 lúnula, 10 sac d’ous, 11 primer pereiopodi, 12 cinquè pereiopodi. 13 tercer pereiopodi, 14 quart pereiopodi, 15 segment genital, 16 turca, 17 tercer segment del perèion.
Gustavo Hormiga, a partir d’originals diversos.
Els copèpodes de vida lliure són crustacis sense closca. La cutícula, a la regió del cap, es diferencia fent un escut cefàlic que únicament es projecta cap a la part anterior ventral i forma un petit rostre, de vegades bifurcat.
La divisió del cos és un dels caràcters peculiars dels copèpodes. L’articulació que separa la part anterior de la posterior permet el moviment dorsiventral de l’una respecte de l’altra, i també una certa mobilitat lateral, particularment important a les espècies nedadores (com els calanoides i moltes espècies de ciclopoides), a les quals la part posterior, clarament més estreta que l’anterior, fa la funció de timó. A les espècies bentòniques o intersticials (com els harpacticoides i alguns ciclopoides), l’amplada del cos és molt més uniforme i la part posterior és poc mòbil. El punt d’articulació de les dues regions del cos no sempre es troba entre els mateixos segments. Segons aquest caràcter, es distingeixen dos grans grups dins dels copèpodes: el dels gimnoplèons, als quals l’articulació separa els segments toràcics (proveïts d’apèndixs) dels abdominals (sense apèndixs) i que recull el grup dels calanoides; i el dels podoplèons (que engloba tota la resta de copèpodes), als quals el punt d’articulació es troba entre el quart i el cinquè metàmers toràcics, de manera que la part posterior del cos porta apèndixs (el cinquè parell toràcic a les femelles i el sisè, rudimentari, als mascles). D’altra banda, els metàmers tendeixen a fusionar-se de dues maneres: pel procés de cefalització, els segments toràcics tendeixen a unir-se als cefàlics, mentre que els abdominals tendeixen a fusionar-se entre ells formant el segment o complex genital. A les espècies paràsites la fusió de segments és molt accentuada, de manera que la segmentació es perd, mentre que unes regions s’atrofien i se n’hipertrofien d’altres. Tota aquesta gamma de variació de la metarnerització dels copèpodes ha portat a diferents interpretacions, i a una nomenclatura variada. Segons alguns autors, la part més anterior del cos, formada pel cap i per un o dos metàmers toràcics fusionats, és el cefalosoma, mentre que els metàmers que queden lliures fins al punt d’articulació amb la part posterior del cos constitueixen el metasoma; la regió posterior a l’articulació és l’urosoma. Això no obstant, és més generalitzada la tendència a considerar el cos dels copèpodes dividit en només dues regions separades per una articulació: el metasoma i l’urosoma.
Els copèpodes porten sis parells d’apèndixs cefàlies (antènules, antenes, mandíbules, maxíl·lules, maxil·les i maxil·lípedes) i 5 o 6 parells d’apèndixs toràcics. Els segments corresponents a l’abdomen no porten apèndixs.
Les antènules, unirràmies i multiarticulades, són els apèndixs més desenvolupats de tot el cos, tot i que varien molt segons que es tracti d’espècies nedadores o bentòniques: a les nedadores, són llargues i fetes de nombrosos segments (de 20 a 25 en els calanoides i de fins a 18 en els ciclopoides), mentre que a les bentòniques són curtes i fetes de pocs segments (3 en el cas d’alguns harpacticoides). No solament són l’òrgan locotomotor principal, sinó que també tenen funció sensorial, perquè porten un gran nombre de sedes al llarg de tots els segments. Als mascles, una de les antènules (en el cas d’alguns calanoides) o totes dues (cas dels ciclopoides i harpacticoides) són geniculades, cosa que els permet subjectar la femella durant la còpula. Les antenes són molt més reduïdes; són birràmies a les formes més primitives (els calanoides i harpacticoides) i unirràmies per la pèrdua de l’exopodi a les més evolucionades (els ciclopoides). Les mandíbules generalment són bífides, però moltes vegades perden l’exopodi; tenen el basipodi molt desenvolupat i quitinitzat, mentre que les dues branques són reduïdes i tenen una estructura semblant a la dels palps mandibulars d’altres grups de crustacis. Les maxíl·lules són birràmies, mentre que les maxil·les i els maxil·lípedes tenen una sola branca. L’estructura d’aquests tres parells d’apèndixs varia segons el tipus d’alimentació; a les espècies lliures responen a dues adaptacions extremes: la microfàgia i la macrofàgia. A les espècies micròfagues (filtradores i detritívores), els apèndixs bucals van proveïts de sedes nombroses a les vores dels diferents artells i, en conjunt, formen els filtres que, moguts com a ventalls, produeixen els corrents d’aigua que porten les partícules alimentàries cap a la boca. Els macròfags han perdut bona part de les sedes dels artells mentre que d’altres s’han transformat en espines, cosa que permet utilitzar aquests apèndixs com a garres prensores, llargues i robustes, per a subjectar les preses.
Els apèndixs toràcics se situen a continuació dels maxil·lípedes. Els quatre primers parells són principalment locomotors; cadascun és format per una coxa, un basipodi i dues branques (l’endopodi i l’exopodi) triarticulades i proveïdes de nombroses sedes i espines; la placa intercoxal uneix els dos apèndixs d’un mateix parell i fa que ambdós es moguin alhora. El cinquè parell d’apèndixs, en general, és molt més reduït i sol presentar dimorfisme sexual. En els calanoides, però, són ben desenvolupats i en els mascles sovint són asimètrics: el dret acostuma a ser molt més llarg i acaba fent una garra, amb la qual subjecta la femella, mentre que l’esquerre acaba en una pinça, que serveix per a transferir l’espermatòfor; paral·lelament, el metàmer corresponent a aquest darrer parell de potes mostra també una lleugera asimetria que dona una petita curvatura a l’eix longitudinal del cos. En els ciclòpides i els harpacticoides, el cinquè parell d’apèndixs és molt més reduït, simètric i semblant en ambdós sexes; com en el cas dels calanoides, aquests apèndixs es relacionen amb la reproducció, tot i que són molt menys especialitzats i mai no serveixen per a subjectar la femella durant la còpula. La utilització del cinquè parell d’apèndixs per a la reproducció fa que tinguin una gran especificitat d’estructura, i per això aquesta constitueix un caràcter important per a la taxonomia del grup. En els ciclopoides s’observa una tendència a la reducció del nombre d’artells, de sedes i d’espines; als gèneres més primitius, com Macrocyclops, cadascun d’aquests apèndixs porta tres artells, mentre que als més modificats com Ectocyclops, es troben molt reduïts i únicament es manifesten en la presència de tres petites espines implantades damunt del tegument. Els mascles dels ciclopoides i dels harpacticoides, i de vegades també les femelles, poden tenir un sisè parell d’apèndixs molt reduït.
A l’urosoma, la tendència dels metàmers a fusionar-se es manifesta en la formacio del complex o segment genital. A les formes lliures són dos els apèndixs abdominals que formen aquest segment, perfectament diferenciat de la resta de metàmers pel fet d’ésser més ample i més llarg. L’interior del complex genital és ocupat pel receptacle seminal a les femelles, i per la vesícula seminal en els mascles. Fins a aquesta cavitat arriben els gonoductes que vénen de la part anterior del cos, i d’ella surten un (cas dels calanoides i harpacticoides) o dos (cas dels ciclopoides i alguns harpacticoides) vasos deferents que desemboquen en un nombre igual de gonopors. L’urosoma acaba en una furca caudal amb dues branques, proveïda de sedes, molt ben desenvolupada a les espècies nedadores i més reduïda a les bentòniques i les intersticials.
Espècies paràsites
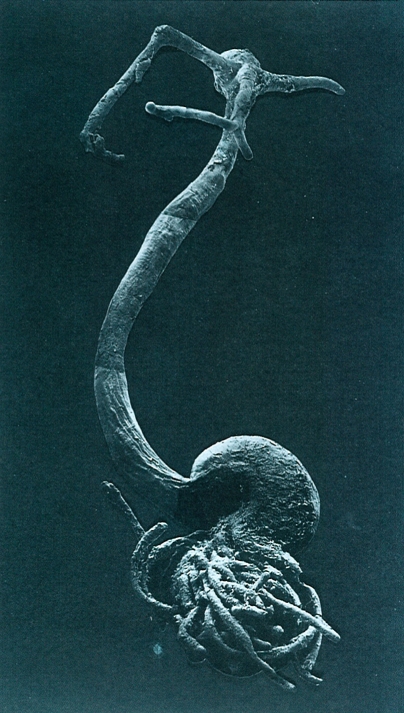
L’aspecte extern dels copèpodes paràsits és extraordinàriament divers i, sobretot, molt modificat respecte d’allò que caracteritza les espècies de vida lliure, especialment en les femelles. A la fotografia, d’una femella de Lernaea branchialis, paràsit de peixos, hom pot distingir les prolongacions cefàliques per mitjà de les quals es fixa a l’hoste, i els cordons ovígers del final del cos.
Montserrat Poquet.
Les modificacions morfològiques que afecten les espècies paràsites són molt variables, i depenen tant del grau d’adaptació a la vida parasitària com de les diferències pròpies de cada grup. En el cas dels harpacticoides, les poques espècies ectoparàsites que se’n coneixen no es diferencien de les formes lliures més que pel tipus de vida. En canvi, en els ciclopoides paràsits (agrupats dins la família dels lernèids), les modificacions són més profundes, però afecten exclusivament les femelles, ja que els mascles no són paràsits. Els monstril·loides representen un cas especial dins dels copèpodes pel fet de tenir les formes larvàries paràsites i les adultes lliures i mancades de boca i de tub digestiu. Finalment, els sifonostomatoides i els pecilostomatoides mostren un grau de modificació molt variable segons cada espècie, però el tipus d’organització general de la boca i els apèndixs bucals dins de cadascun d’aquests grups respon a un mateix model.
Aquest conjunt de modificacions es pot resumir en una sèrie de tendències que afecten tant els metàmers com els apèndixs. D’una banda, s’observa una tendència a perdre la segmentació del cos, cosa que ja es manifesta a les espècies de vida lliure. Aquesta indiferenciació del cos s’acompanya de l’atròfia dels apèndixs locomotors, que fins i tot arriben a desaparèixer. Paral·lelament, apareixen estructures relacionades amb els apèndixs cefàlics dedicades a facilitar la fixació del paràsit damunt del seu hoste. Per exemple, les antènules poden ser molt reduïdes mentre que les antenes, les maxíl·lules, les maxil·les i els maxil·lípedes es modifiquen de maneres diverses per a formar garres, ungles, dents i ventoses que constitueixen els elements de fixació. El labre i el llavi també es modifiquen per a formar, juntament amb les mandíbules, un sifó adaptat a la succió. Tret de les modificacions que afecten la zona bucal, que tenen valor taxonòmic, la resta de modificacions són generals per a tots els ordres.
Paral·lelament a les modificacions de la morfologia externa, s’observen diferències internes importants, respecte de les espècies de vida lliure, per exemple en l’estructura i la ultrastructura del tegument, de la musculatura esquelètica, dels òrgans sensorials i de l’aparell digestiu. D’altra banda, l’aparell reproductor experimenta una hipertròfia considerable, fins al punt que en algunes espècies és l’òrgan més voluminós del cos.
Biologia
Espècies de vida lliure

El dimorfisme sexual sol ésser important dins dels copèpodes. A les espècies de vida lliure aquest es manifesta en l’existència, en els mascles d’antenes geniculades (una sola o totes dues), amb les quals sostenen la femella durant la còpula, i, de vegades, d’una asimetria bilateral del final del cos, que pot ésser més o menys acusada. Hom pot observar aquests caràcters lligats al sexe en els exemplars mascle (a dalt) i femella (a baix) d’un diaptòmid (Hemidiaptomus roubaui), comú a les aigües dolces temporànies del país.
Enric Ribes.
Tret d’un petit nombre d’espècies partenogenètiques (dins dels harpacticoides), tota la resta dels copèpodes es reprodueixen sexualment. Els mascles guarden els espermatozoides en unes càpsules o estoigs anomenades espermatòfors que, posteriorment, durant la còpula, enganxen a la femella, mentre que la mantenen subjecta per mitjà de l’antènula geniculada i el cinquè parell d’apèndixs (en els calanoides) o de les dues antènules (en els ciclopoides i harpacticoides). Una sola còpula és suficient per a fecundar tots els ous d’una femella, però cada femella pot copular successivament amb diversos mascles, i transportar, per tant, diversos espermatòfors. Els espermatozoides no són mòbils i no es coneix amb certesa la manera com surten dels espermatòfors per a fecundar els ous de la femella. A les espècies d’aigües continentals i en algunes espècies marines la mare transporta els ous fecundats dins d’un (en els calanoides i harpacticoides) o dos (en els ciclopoides) sacs ovígers segregats per les glàndules annexes al sistema reproductor. Moltes de les espècies marines abandonen els ous a mesura que els van produint. El nombre d’ous de cada posta varia a cada espècie, en general, en relació amb les dimensions de la mare i del seu nombre de cromosomes; hom creu que altres factors, com la temperatura o el pH, poden influir en les proporcions sexuals de la nova generació.
El desenvolupament dels ous generalment és immediat, però en algunes espècies que viuen a l’aigua dolça, especialment les d’aigües temporànies, poden produir-se uns ous especials, més grans i amb una coberta gruixuda, que són de desenvolupament diferit.
Dels ous neixen larves naupli, el·líptiques i proveïdes de tres parells d’apèndixs (antènules, antenes i mandíbules), i a través de sis estadis nauplians van incorporant les maxíl·lules, les maxil·les, els maxil·lípedes i els dos primers parells d’apèndixs toràcics; en tots aquests estadis no hi ha una segmentació manifesta del cos. A continuació s’inicia la fase de copepodit, que es caracteritza per l’aparició de segmentació i per l’aspecte general del cos, molt semblant al de l’adult; n’hi ha cinc estadis diferents, cadascun dels quals incorpora nous metàmers i nous apèndixs, i a partir del tercer es manifesta ja el dimorfisme sexual. Alguns ciclopoides tenen només cinc estadis nauplians. En resum, les espècies de vida lliure passen per 11 o 12 estadis larvaris, caracteritzats pel nombre i la morfologia dels apèndixs i per la segmentació progressiva del cos. La transformació gradual dels apèndixs, especialment els bucals, suposa canvis importants de l’alimentació i de la modalitat de captura de l’aliment, de manera que cada espècie es podria equiparar a un conjunt d’espècies amb espectres alimentaris diferents.
La durada de la vida dels copèpodes generalment oscil·la entre 3 i 9 mesos, amb variacions que depenen de la biologia de cada espècie. A les aigües continentals, i d’una manera aproximada, es pot dir que les espècies litorals, bentòniques i de petites dimensions són de vida curta (d’1 a 9 mesos) i tenen moltes generacions cada any, mentre que les espècies planctòniques, generalment formes grans, solen viure de 9 a 12 mesos, i no tenen més de dues generacions a l’any. A les espècies de vida curta, les diferents generacions es desenvolupen en condicions ambientals diferents (de temperatura, aliment, etc.), de manera que es diferencien no sols genèticament, sinó també com a resultat de les condicions ambientals ara esmentades; això fa que els caràcters morfomètrics siguin de valor dubtós per a la taxonomia. En canvi, en el cas de les espècies de vida llarga, i més quan es compara entre generacions desenvolupades dins d’una mateixa època de l’any, les diferències de morfometria entre poblacions es poden emprar per a diferenciar subespècies, varietats o races, que reflecteixen el grau d’adaptació a l’ambient i permeten estudiar la diferenciació geogràfica intraspecífica.
Espècies paràsites
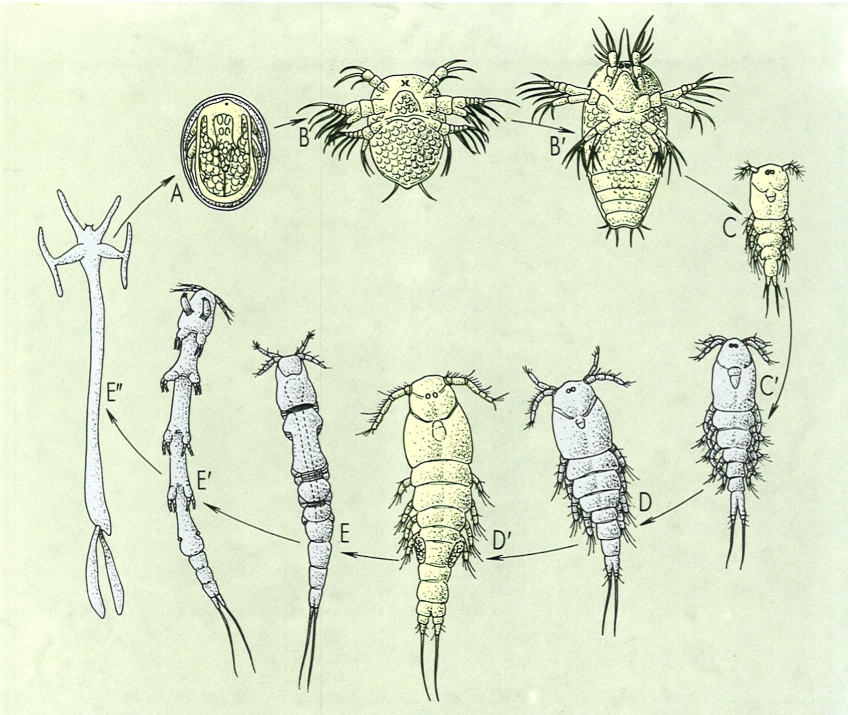
Cicle de vida del copèpode paràsit Lernaea branchialis. Acomplert el desenvolupament embrionari dins de l’ou (A), l’espècie inicia el seu desenvolupament postembrionari en estat de naupli (B, B’), de vida lliure, el qual evoluciona fins a l’estat de copepodit (C, C’), i aquest, després de passar per diverses fases es fixa al damunt d’un peix i hi resta fins a atènyer l’estadi d’adult (D). Les femelles, (D’) després d’ésser fecundades pel mascle, abandonen el primer hoste i van a fixar-se a les brànquies d’un altre peix, pertanyent a una altra espècie. Són les femelles les que experimenten grans modificacions en la seva morfologia (E, E’. E") fins a divergir completament dels copèpodes típics i, profundament enfosades en el cos de l’hoste, creixen fins a atènyer grandàries de 15 i 25 cm.
Servei de Fotografia/Román Montull, a partir de fonts diverses
Els copèpodes paràsits mostren una reproducció de tipus sexual i un cicle de desenvolupament semblant als de vida lliure, bé que amb algunes diferències pròpies de la vida parasitària. A les espècies paràsites el dimorfisme sexual és molt més accentuat, i afecta les dimensions del cos i la forma de vida: en algunes espècies, únicament les femelles són paràsites. Per exemple, els mascles de Nadranchia i de Myzotheridion viuen fixats al damunt del cos de la femella, i són unes 50 vegades més petits que aquesta. Els dos sacs ovígers són molt grossos, de vegades fins i tot més grossos que el cos de la femella, i poden contenir milers d’ous; són ovalats (com al gènere Mytilicola) o cilíndrics (com en Lernanthropus). Per les dificultats de coincidència dels individus adults de sexe contrari, generalment la fecundació es fa durant els últims estadis larvaris.
Dels ous neixen larves naupli, que queden lliures dins dels teixits de l’hoste, a la cambra paleal en el cas dels mol·luscs, o a l’aigua, en el cas de les espècies ectoparàsites. Les fases de naupli i copepodit acostumen a ser de vida lliure i presenten una morfologia semblant a la de les espècies lliures. En atènyer la fase d’adult, els individus (en algunes espècies, les femelles) es fixen a l’hoste, bé que de vegades la fixació ja es produeix durant alguns dels estadis de copepodit, com és el cas de Mytilicola o de Lernaea. En el grup dels monstril·loides, aquest cicle és invertit, de manera que les larves són endoparàsites i els adults són lliures.
Ecologia
Espècies de vida lliure

Els copèpodes constitueixen una part molt important de la biomassa animal en el zooplàncton de les aigües continentals i marines. A la fotografia hom pot veure una mostra de calanoides (de l’espècie Calanus helgolandicus), presa a la zona profunda de la mar Mediterrània, que pot il·lustrar la morfologia típica general dels copèpodes de vida lliure.
Francesc Vives.
Tot i que, comparats amb altres grups d’artròpodes, els copèpodes no són els més diversificats, representen el grup animal més abundant del planeta, en gran part gràcies a les espècies planctòniques, marines i d’aigües continentals. Tant pel nombre d’individus com per la biomassa que representen, els copèpodes planctònics tenen una gran importància a les cadenes tròfiques dels ecosistemes als quals es troben. Generalment no són gaire estrictes pel que fa al tipus d’alimentació (herbívora, detritívora o carnívora), atès que el canvien al llarg del seu desenvolupament. Per això, potser resulta més idoni parlar de microfàgia i macrofàgia, aquesta més pròpia dels estadis finals de desenvolupament de les formes més grans. Fins i tot en les espècies estrictament filtradores, la filtració no és passiva, en funció de la separació de les sedes, sinó que són capaces de detectar la presència de partícules d’aliment per mitjà de quimioceptors o mecanoceptors, i de moure els apèndixs bucals per a capturar-les d’una manera selectiva. D’altra banda, els copèpodes formen part de la dieta d’un gran nombre d’organismes marins, especialment planctònics (meduses, ctenòfors, quetognats, peixos) o d’aigües continentals (larves de Chaoborus, Mysis, peixos).
Al mar, els copèpodes pelàgics, predominantment clanoides, són els més abundants i coneguts. A la Mediterrània n’hi ha unes 280 espècies pelàgiques, majoritàriament d’origen atlàntic, de les quals es troben aproximadament la meitat a la zona occidental. La diversitat de les poblacions augmenta amb la fondària i la distància a la costa, mentre que el nombre d’individus disminueix. Per tant, als 50 m superficials s’acumula el 78 % dels individus, entre els 50 i els 200 m el 16 %, entre els 200 i els 500 m el 4 % i entre els 500 i els 1000 m el 2 % restant.

L’abundància i la distribució dels copèpodes planctònics marins és avaluada per mitjà de diverses tècniques fonamentades en el mostratge selectiu d’àrees i fondàries amb xarxes especials. Un exemple de l’aplicació d’aquestes tècniques d’estudi és aquesta col·lecció de mostres preses a diferents fondàries, en una àrea situada davant de l’aeroport del Prat (Barcelona), que demostra d’una manera ostensible l’heterogeneïtat vertical del poblament bentònic marí. De dreta a esquerra, les provetes 73 i 74 contenen mostres preses a 12 m de fondària, constituïdes gairebé exclusivament per Sagitta setosa, un quetognat. Les provetes 72 i 71, procedents de 25 m de fondària, contenen S. setosa i Paracalanus parvus i Temora stylifera, dos copèpodes. Les provetes 70 i 69, procedents d’una àrea de 50 m de fondària, contenen anomurs, decàpodes, i copèpodes de diferents gèneres i espècies.
Francesc Vives.
A aquesta distribució vertical se superposa una sèrie de canvis temporals, que afecten els 200 m superficials. A l’estiu, quan l’aigua es troba estratificada (a causa del gradient de densitat originat per l’escalfament de les capes superficials de l’aigua), les poblacions queden desplaçades cap al fons i se situen preferentment entre els 50 i els 100 m, coincidint amb concentracions màximes de clorofil·la. En una escala de temps més curta, són importants les migracions verticals de ritme diari (nictamerals), que afecten principalment els copèpodes situats als primers 200 m. Al capvespre pugen a la superfície a partir del punt on passen el dia, a mitjanit fan un petit descens, per a tornar a pujar a l’alba; a primeres hores del matí baixen fins a la posició inicial del cicle. El significat d’aquest fenomen no és encara ben conegut; de fet, és sorprenent, si es pensa en la quantitat d’energia que cal invertir per a fer migracions de fins a 200 m, tenint en compte que es tracta d’animalets d’uns 2 mm de llargada. El que és certés que aquests moviments es relacionen amb ritmes alimentaris, acoblats a un factor de regulació, com és el canvi d’intensitat de llum al llarg del dia. Les conseqüències d’aquests comportaments són importants dins la dinàmica de tot l’ecosistema en conjunt, ja que representen un transport accelerat de matèria orgànica des de la superfície (on els copèpodes s’alimenten) fins a les zones profundes; d’altra banda, el fet que els excrements dels copèpodes vagin compactats i embolicats amb una membrana peritròfica ("fecal pellets") incrementa considerablement la seva velocitat de sedimentació.
Les comunitats pelàgiques de la Mediterrània es poden separar en tres grups: les de les zones marines confinades (badies, ports, estuaris), les nerítiques i les batipelàgiques. Dins del primer grup s’observa una predomináncia dels calanoides de la família dels etidèids, amb grups d’espècies congenèriques que són representants típics de zones amb una elevada variabilitat ambiental una certa desalinització o contaminació (Acartia clausi, A. grani, A. margalefi, A. discaudata var. mediterranea, A. latisetosa); també dins d’aquest grup hi ha algunes espècies de ciclopoides de la família dels oitònids (com Oithona nana i O. helgolandica) i els harpacticoides Euterpina acutifrons, Clitemnestra scutellata i algunes espècies del gènere Tisbe; altres espècies, com Temor a stylifera, Centropages typicus, Paracalanus parvus i Oncaea media, provenen de la zona nerítica, i la seva presència en aquests ambients és més aviat estacional. A la zona nerítica hi ha al voltant d’unes 80 espècies de copèpodes, però d’aquestes, només una quinzena constitueixen el 90 % dels individus. A més d’algunes espècies dels gèneres Acartia i Oithona ja esmentades per a les zones semiconfinades, hi són freqüents Centropages typicus, Temora stylifera, Clausocalanus, Ctenocalanus vanus, Paracalanus parvus i Oncaea media; també s’hi afegeixen d’altres formes que generalment viuen a zones més profundes, i que arriben als nivells superiors quan les condicions hidrogràfiques els són propícies (a l’hivern i a començament de primavera, que és quan no hi ha gradient vertical de densitat, perquè la temperatura de l’aigua és la mateixa a tot el perfil), com Calanus helgolandicus, tal vegada l’espècie de copèpode més estudiada, i espècies no tan freqüents, dels gèneres Aetideus, Euchaeta i Pleuromamma. Finalment, a la zona batipelàgica, per sota dels 200 m, s’observa una gran uniformitat pel que fa a la distribució d’espècies, representades principalment per les famílies dels etidèids (Euaetideus giesbrechti, Aetideus armatus), els euquètids (Euchaeta marina, E. curta), els escolecítrids (Scaphocalanus curtus, Scolecithricella abissalis, Pleuromamma abdominalis), els heteroràbdids (Heterorhabdus papilliger) i els oncèids (Oncaea curta); la diversitat d’espècies hi és, doncs, elevada, cosa que contrasta amb una poca abundància en nombre d’individus a tota la zona.
Els copèpodes bentònics, comparats amb els planctònics, són molt menys coneguts. El seu interès des del punt de vista de les xarxes tròfiques es redueix a la zona litoral, on exploten com a micròfags detritívors la part superficial del sediment, o bé formen part de la meiofauna. En aquesta zona es troben, gairebé d’una manera exclusiva, espècies d’harpacticoides i algun ciclopoíde. A les zones fangoses dels estuaris són comunes Tachidius discipes, Nannopus palustris (espècie que també es troba a les aigües continentals) i Paronychocamptus nanus. Associades a substrats amb algues, hi ha Harpacticus obscurus, Parathalestris harpacticoides i Dactylopodia vulgaris, entre moltes altres espècies no tan freqüents; per contra, Tisbe ensifer i Metis ignea són molt més indiferents al substrat i, per tant, mostren una distribució més àmplia.
Les aigües litorals salabroses, sigui intersticials o epicontinentals, constitueixen un bon exemple del fenomen d’invasió per copèpodes marins. Les aigües intersticials del litoral (platges, desembocadures de rius, etc.) han estat poc estudiades, però pel que sembla s’hi troben copèpodes, principalment harpacticoides, amb espècies principalment marines, o bé amb formes molt pròximes a espècies marines. Per exemple, a la desembocadura del riu Ter, entre l’aigua intersticial litoral, s’hi troben, entre altres, Parastenocaris, Psammastacus confluens i Robetgurmeya. A les llacunes litorals d’aigües salabroses, la composició específica varia molt segons la salinitat: al costat d’espècies marines, com Harpacticus littoralis i Metis ignea, n’apareixen d’altres més resistentes a les variacions de salinitat, com Nitocra lacustris, i fins i tot Bryocamptus pigmaeus, que generalment viu en aigües poc salabroses.
La colonització de les aigües continentals per calanoides és representada a les nostres costes per tres grups d’espècies que mostren nivells d’adaptació creixent. Dins d’un primer grup se situen les espècies marines que poden viure a les aigües confinades que reben alguna font d’aigües continentals, com és el cas d’algunes espècies d’Acartia: A. latisetosa, del delta de l’Ebre, l’albufera de Maó, el port de Barcelona; A. margalefi, trobada als canals d’Empuriabrava. El segon grup inclou les espècies típicament litorals, que no es troben ni a la mar ni a les aigües continentals poc salades, com és el cas de Calanipeda aquae-dulcis. El tercer grup reuneix espècies del gènere Eurytemora, que mostren un grau d’adaptació més alt a les aigües de poca salinitat, i de les quals únicament coneixem E. velox, als aiguamolls de l’Empordà.
A les aigües continentals, com a la mar, els copèpodes constitueixen un grup animal important, bé que amb menys espècies. Per exemple, a la península Ibèrica se’n coneixen un centenar d’espècies, de les quals unes 70 són presents als Països Catalans. Diferent del que passa a les comunitats marines, al plàncton de qualsevol llac o embassament sol haver-hi 3 o 4 espècies, i sovint només una o dues. En canvi, la discontinuïtat ambiental pròpia d’aquests hàbitats permet una diversificació molt més gran de les comunitats.
La coexistència de diferents espècies de copèpodes implica l’adopció d’estratègies alimentàries diferents. Per exemple, al llac de Banyoles, entre el plàncton hi ha el calanoide filtrador (micròfag) Arctodiaptomus salinus, dos ciclopoides herbívors (macròfags) de grandària diferent (Tropocyclops prasinus i Diacyclops bicuspidatus), i un ciclopoide carnívor (macròfag), Cyclops abyssorum. Als llacs dels Pirineus es troben comunitats molt més variades, d’acord amb les característiques de cada llac. En els casos més senzills hi ha una sola espècie de copèpode (Cyclops abyssorum o Acanthocyclops vernalis), mentre que a les comunitats més complexes pot haver-hi dos calanoides de dimensions diferents (Diaptomus cyaneus amb Mixodiaptomus laciniatus o Eudiaptomus vulgaris) i dos ciclòpids, dels quals un és herbívor i l’altre és carnívor.
Als embassaments, les comunitats mostren unes característiques semblants a les dels llacs, però mantenen més variabilitat, per dues raons: per la seva construcció recent i pel fet d’ésser ecosistemes molt fluctuants. En general es pot dir que a tota la regió mediterrània hi ha un grup de copèpodes de distribució molt general, amb una gran capacitat d’adaptació a tota mena de condicions ambientals, format per Tropocyclops prasinus, diverses espècies d’Eucyclops (E. serrulatus, E. speratus), Macrocyclops albidus i Acanthocyclops (A. venalis, A. robustus). A la península Ibèrica, l’escassesa de llacs grans ha limitat molt la presència d’espècies planctòniques, que són molt comunes a la zona temperada (especialment del gènere Cyclops) o tropical (Mesocyclops, Thermocyclops, Diacyclops).
A les aigües continentals, els diaptòmids són els representants més conspicus dels copèpodes calanoides. En general són força selectius respecte de les condicions ambientals, i això fa que entre ells hi hagi espècies indicadores; per exemple, Neolovenula alluaudi viu en aigües riques en sulfats, pròpies de zones endorreiques, amb guixos abundants; Arctodiaptomus (A. salinus i A. wierzejskii) viu en aigües atalassohalines; Copidodiaptomus numidicus i Diaptomus castaneti a les aigües poc mineralitzades eutròfiques i oligotròfiques, respectivament. Les aigües temporànies també tenen representants característics entre els diaptòmids, bé que, pel fet de trobar-se menys segregats, és freqüent de trobar-hi fins a tres espècies juntes (Hemidiaptomus roubaui, Diaptomus cyaneus i Mixodiaptomus incrassatus), sovint de dimensions diferents. A més de les espècies esmentades, hi ha petits copèpodes (de menys d’un mm), poc adaptats a la natació, que viuen entre la vegetació o al sediment; dins d’aquest grup hi ha algunes espècies petites de ciclopoides (com Paracyclops fimbriatus, Metacyclops minutus, Ectocyclops phaleratus) i la gran majoria dels harpacticoides, els quals, malgrat trobar-s’hi molt diversificats, són poc coneguts. Únicament Canthocamptus staphylinus apareix esporàdicament entre les mostres de plàncton.
Espècies paràsites
Dins dels copèpodes es presenten totes les modalitats de vida parasitària: ectoparasitisme, endoparasitisme, hiperparasitisme, fases larvàries lliures que es transformen en adults paràsits i viceversa, paràsits d’alta i baixa especificitat d’hoste, etc.
En general, les modificacions morfològiques són un bon indicador del grau d’integració de cada espècie a la vida paràsita. Per exemple, alguns harpacticoides i ciclopoides paràsits únicament difereixen de les formes lliures pel seu tipus de vida; i, al contrari, en el cas dels endoparàsits, les modificacions són tan profundes que únicament es poden reconèixer com a copèpodes en alguns estadis del seu cicle biològic.
La majoria de les espècies paràsites són marines, i és en aquest ambient on han assolit la màxima diversificació: arriben a parasitar gairebé tots els grups marins de metazous (equinoderms, ascidis, mol·luscs, anèl·lids, peixos, cetacis). En canvi, a les aigües continentals, el nombre d’espècies paràsites és reduït.
A les modificacions pròpies del parasitisme, cal afegir-hi, en molts casos, un dimorfisme sexual molt accentuat. En algunes espècies és només la femella qui porta una vida parasitària, com el cas de Ergasilus sieboldi, que viu a la llacuna de la Ricarda (delta del Llobregat); en d’altres, com dins de la família dels condracàntids, el mascle és molt petit i roman fixat a la superfície de la femella, prop de l’orifici genital (per exemple, Chondracanthus merlucci, de la qual el mascle és 12 000 vegades més petit que la femella).
L’especificitat de les relacions entre el paràsit i l’hoste és un bon indicador del grau de coevolució assolit. Per exemple, el llobarro (Dicentrarchus labrax) pot actuar d’hoste, simultàniament, de Caligus minimus i Lernanthropus kroyerii; Notobranchis syquiformis viu sobre la boga (Boops boops), l’esparrall (Diplodus annularis), el sard (Diplodus sargus), el pagre (Sparus pagrus) i la xucla (Smaris smaris). També és freqüent que una mateixa espècie pugui parasitar diferents hostes successivament al llarg del seu cicle biològic i, en aquest cas, els cicles van sincronitzats amb les relacions tròfiques del sistema, ja que depenen de la depredació per a transmetre’s des d’un animal que actua de presa fins al predador; el cicle de Lernaeocera branchialis és un bon exemple: després d’una fase inicial lliure (naupli, metanaupli i copepodit), l’individu es fixa a les brànquies del llenguado (Solea vulgaris) on completa el seu desenvolupament i té lloc la reproducció; posteriorment, les femelles abandonen l’hoste i van a fixar-se a les brànquies d’un bacallà (Gadus macrocephalus), on creixen extraordinàriament, tant que poden arribar fins al cor del peix. En els casos que el cicle es fa al llarg d’una cadena tròfica, el paràsit sol influir sobre la conducta i el color de l’hoste per tal que aquest li faciliti el seu trasllat a un altre hoste. En el cas dels musclos (Mytilus) parasitats per Mytilicola intestinalis, en el seu hepatopàncrees verdós es poden distingir, per llur color vermellós, de 6 a 8 individus de Mytilicola d’ambdós sexes. Un altre efecte dels copèpodes paràsits sobre els seus hostes és l’esterilització: el paràsit Mytilicola, quan envaeix els teixits dels musclos (Mytilus edulis i M. galloprovincialis), pot arribar fins a les gònades i aprofitar-ne les reserves destinades a la reproducció, mentre produeix l’esterilitizació de l’individu. El cas de Mytilicola intestinalis és un exemple d’hiperparasitisme, ja que aquesta espècie paràsita de musclos pot ésser, al seu torn, parasitada per un esporozou (Unicaryon mytilicolae), que viu als seus teixits somàtics i en molts casos també es troba en els gàmetes, per la qual cosa els ous embrionats dels sacs ovígers ja tenen el paràsit.
Sistemàtica
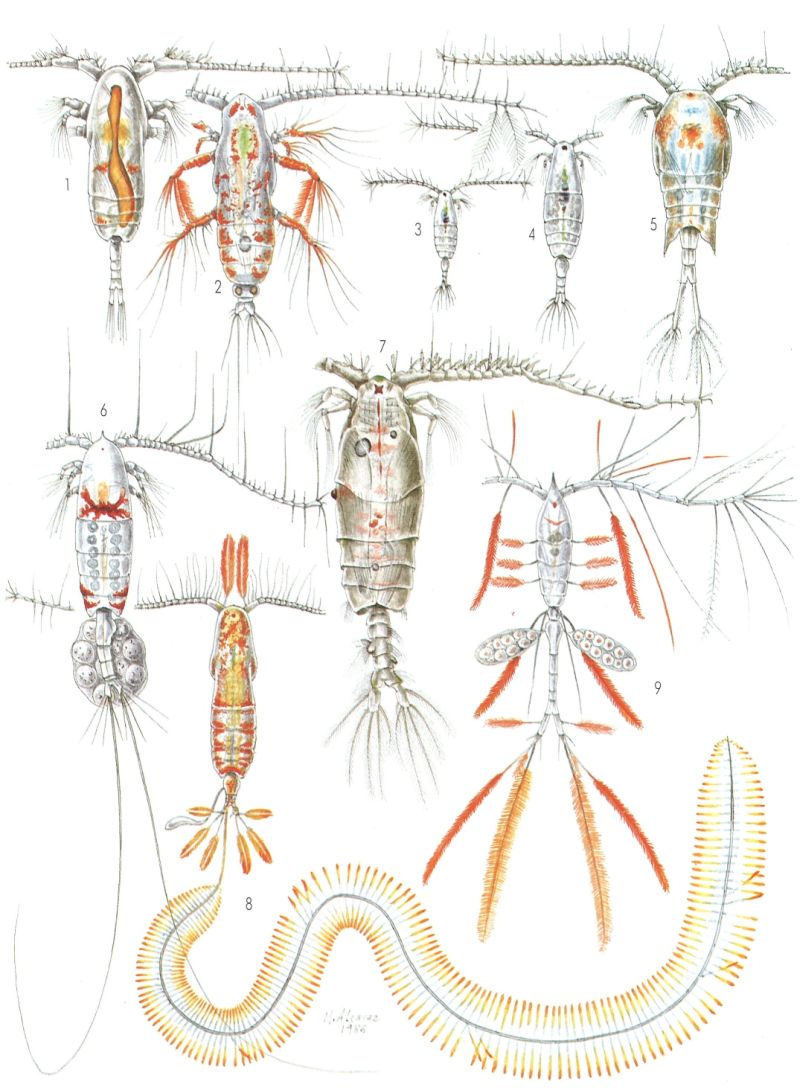
Copèpodes marins, I. 1 Paracalanus parvus, mascle (× 60). 2 Eucalanus crassus, femella (× 17). 3 Acartia margalefl, femella (× 30). 4 A. clausi, femella (× 30). 5 Temora stylifera, femella (× 33). 6 Euchaeta marina, femella (× 20). 7 Pleuromamma abdominalis, mascle (× 23). 8 Calocalanus plumulosus, femella (× 50). 9 Oithona plumifera, femella (× 47).
Miquel Alcaraz.

Copèpodes marins, II. 1 Candacia aethiopica, femella (× 20). 2 Calanus helgolandicus, femella (× 27). 3 Sapphirina ovatolanceolata, mascle (× 17). 4 Anomalocera patersoni, mascle (× 20). 5 Centropages typicus, femella (× 37). 6 Oncaea venusta, femella (× 47). 7 Oithona nana, femella ovígera (× 83). 8 Calocalanus pavo, femella (× 47). 9 Corycaeus (Onichocorycaeus) giesbrechti, femella ovígera (× 67).
Miquel Alcaraz.
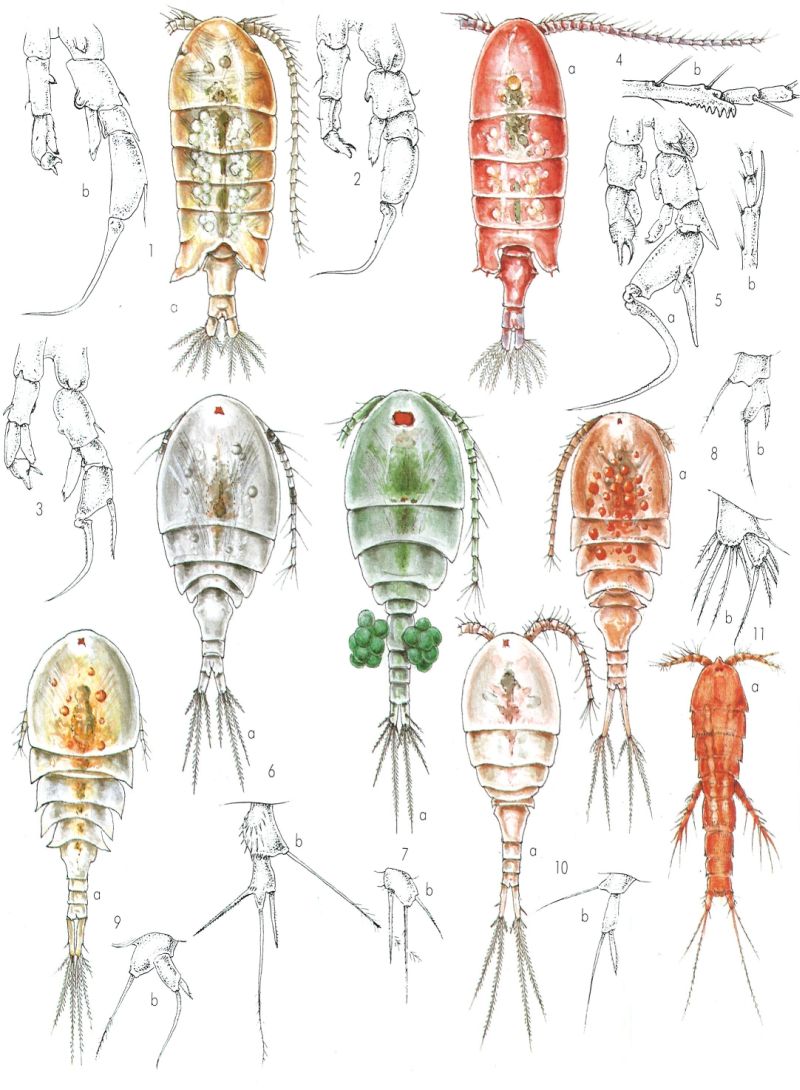
Copèpodes d’aigua dolça. 1 Mixodiaptomus kupelwieseri: a femella (× 33), b cinquè apèndix toràcic d’un mascle. 2 M. incrassatus, cinquè apèndix toràcic d’un mascle. 3 M. laciniatus, cinquè apèndix toràcic d’un mascle. 4 Arctodiaptomus wierzejskii: a femella (× 30), b tres darrers segments de l’antena d’un mascle. 5 A. salinus: a cinquè apèndix toràcic d’un mascle, b tres darrers segments de l’antena d’un mascle. 6 Macrocyclops albidus: a femella (× 30), b cinquè apèndix toràcic d’un mascle. 7 Tropocyclops prasinus: a femella (× 93), b cinquè apèndix toràcic d’una femella. 8 Cyclops abyssorum mauritaniae: a femella (× 40), b cinquè apèndix toràcic d’una femella. 9 Acanthocyclops vernalis: a femella (× 57), b cinquè apèndix toràcic d’una femella. 10 Diacyclops bicuspidatus: a femella (× 60), b cinquè apèndix toràcic d’una femella. 11 Canthocamptus staphylinus microstaphylinus: a femella (× 80), b cinquè apèndix toràcic d’una femella.
Miquel Alcaraz.
Els copèpodes (Copepoda) constitueixen una subclasse de la classe dels crustacis, la sistemàtica dels quals ha estat alterada recentment gràcies a la nova informació que han donat els estudis fets sobre grups paràsits; efectivament, fins no fa gaire tota la sistemàtica es fonamentava en l’analogia dels caràcters morfològics entre els diferents grups, sense tenir en compte que aquestes analogies moltes vegades són resultat de l’adaptació a la vida parasitària. El sistema de classificació de Kabata, que és el que s’utilitza ací, s’estructura sobre criteris que separen clarament els caràcters anàlegs dels homòlegs. Segons aquest sistema, s’accepta que la subclasse dels copèpodes inclou vuit ordres, set dels quals són representats al nostre país: calanoides (Calanoida), harpacticoides (Harpacticoida), ciclopoides (Cyclopoida), pecilostomatoides (Poecylostomatoida), sifonostomatoides (Sifonostomatoida), monstril·loides (Monstrilloida) i mormonil·loides (Mormonilloida). L’ordre dels misofrioides (Misofrioida), comprèn una sola espècie, pròpia de l’oceà Atlàntic, amb la característica peculiar de tenir cor, cosa que la converteix en un cas únic dins dels copèpodes. Tots vuit ordres, en conjunt, reuneixen 8500 espècies.
Els grups de copèpodes
Els calanoides
Els calanoides es consideren el grup amb categoria d’ordre més primitiu de tots els copèpodes actuals, atès que reuneix tots els gimnoplèons. Els seus representants són planctònics, marins i d’aigües continentals. Es caracteritzen per tenir les antenes, principals elements impulsors, formades per un nombre de segments que oscil·la entre 20 i 25. Els calanoides comprenen nombroses famílies, com les dels diaptòmids (Diaptomidae), etidèids (Aetideidae), mitilicòlids (Mytilicolidae), els oncèids (Oncaeidae), dels quals ja hem parlat a bastament.
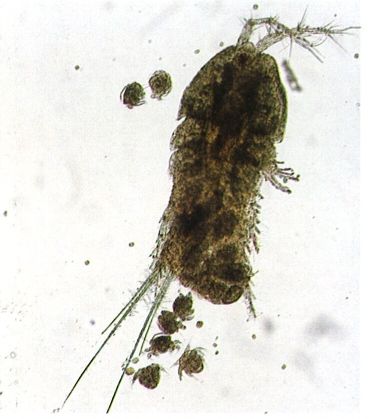
Femella d’un harpacticoide mostrant l’eclosió dels ous i la sortida de les larves naupli. La fotografia, feta a partir d’una mostra presa al port de Barcelona, correspon al gènere Tisbe, bentònic i distribuït per les mars de tot el món, ja que pot viure en aigües de salinitats molt diverses.
Mercè Durfort.

Un calanoide força representatiu és el de la fotografia, Acartia tonsa (un mascle), propi del plàncton superficial de la Mediterrània, com moltes altres espècies del seu mateix gènere.
Mercè Durfort/Xavier Vidal.
Els harpacticoides
Els harpacticoides inclouen la majoria de les espècies de copèpodes bentònics i d’aigües intersticials. El cos cilíndric, les antènules reduïdes i els apèndixs locomotors poc adaptats a la natació són caràcters que defineixen aquests copèpodes que fan vida prop del substrat. Aquest és l’ordre de copèpodes més diversificat: reuneix unes 2800 espècies, de les quals, aproximadament, el 90 % són marines i pertanyen majoritàriament a les famílies dels harpacticídids (Harpacticidae, gènere Harpacticus), mètids (Metiidae, gènere Metis) i tísbids (Tisbiidae, gènere Tisbe). Més de la meitat de les espècies d’aigües continentals pertanyen a la família dels cantocàmptids (Canthocamptidae), dins la qual destaquen per la seva freqüència Canthocamptus i Bryocamptus. D’entre les espècies de les aigües intersticials, destaquen les de la família dels parastenocàrids (Parastenocari-. dae, gènere Parastenocaris), moltes de les qual són endèmiques i d’un gran interès biogeogràfic; es tracta d’un grup d’origen marí que actualment colonitza el medi hipogeu continental.
Els ciclopoides
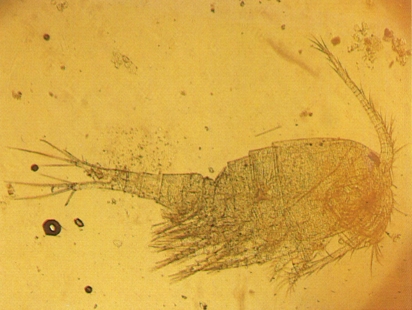
Els ciclòpids són abundants i molt ben diversificats a les aigües continentals. A l’exemplar de la fotografia, fixat i vist al microscopi òptic, pertanyent al gènere Cyclops, hom pot apreciar la diferència d’amplada entre el perèion i el plèon que caracteritza els ciclopoides nedadors.
Víctor Ribera.
Els ciclopoides són espècies principalment lliures, marines i d’aigües continentals. Tenen les antènules amb 10-18 artells i el cos més o menys modificat segons el tipus de vida: a les espècies nedadores, l’urosoma és llarg i estret respecte del metasoma, mentre que a les bentòniques és menys diferenciat. És un grup molt diversificat a les aigües continentals, on fan un paper semblant al que els calanoides fan al medi marí. Les espècies d’aigües continentals pertanyen principalment a la família dels ciclòpids (Cyclopidae, a la qual pertanyen Cyclops, Tropocyclops, Acanthocyclops) i les marines a les famílies dels oitònids (Oithonidae, gènere Oithona) i els oncèids (Oncaeidae, gènere Oncaea), entre d’altres. La família dels lernèids (Lernaeidae) reuneix espècies paràsites (gènere Lernaea).
Els petilostomatoides
Els pecilostomatoides són paràsits d’invertebrats i d’alguns peixos marins. Morfològicament són molt modificats respecte de la morfologia general dels copèpodes, principalment per la pèrdua de la segmentació (a partir de l’estadi de copepodit), l’atròfia de les antènules i la transformació dels altres apèndixs cefàlies en garres, ungles o dents per a subjectar-se a l’hoste i raspar-ne els epitelis. Aquest ordre inclou algunes de les famílies amb espècies molt comunes a les nostres costes, com la dels condracàntids (Chondracanthidae, gènere Chondracanthus), la dels mitilicòlids (Mytilicolidae, gènere Mytilicola·) i la dels ergasílids (Ergasilidae, gènere Ergasilus).
Els sifonostomatoides
Els sifonostomatoides són paràsits de peixos marins i d’aigua dolça, i també d’invertebrats. Com en el grup anterior, moltes de les seves espècies han perdut secundàriament la segmentació del cos i tenen els apèndixs cefàlics modificats per a la fixació. La principal característica que els defineix és la prolongació i fusió del llavi i el labre formant un tub o sifó a l’interior del qual hi ha les mandíbules. És el grup de paràsits més diversificat a la Mediterrània occidental, on se’n troben moltes famílies; les que inclouen les espècies més freqüents són les dels calígids (Caligidae, gènere Caligus), la dels lernantròpids (Lernanthropidae, gènere Lernanthropus), la dels lerneopòdids (Lernaeopodidae, gèneres Neobranquia, Brachiella, Salmicola), els pennèl·lids (Pennellidae, gènere Pennella, Lernaescolea), i la dels herpilòbids (Herpyllobiidae, gèneres Xenocoeloma, Rhizoshina, Herpyllobius), aquesta exclusivament paràsita de poliquets.
Els mostril·loides
Els monstril·loides constitueixen l’únic ordre de copèpodes al qual les formes paràsites es presenten durant els primers estadis de desenvolupament. Com a conseqüència d’aquest cicle biològic, els adults no tenen més apèndixs cefàlics que les antènules, i tenen el tub digestiu atrofiat. Els gèneres Monstrilla, Haemocera, Cymbasoma i Monstrillopsis, paràsits d’oligoquets i gasteròpodes, reuneixen les espècies trobades fins ara al nostre litoral.
Els mormonil·loides
Els mormonil·loides tenen un aspecte intermedi entre els calanoides i els ciclopoides, encara que no tenen els cinc primers parells d’apèndixs toràcics. Les antenes són llargues, però fetes solament per 3 o 4 artells. Se’n coneix un sol gènere (Mormonilla), present a la Mediterrània.