És un rosegador de talla mitjana amb les potes anteriors i posteriors no gaire llargues en proporció amb el cos. Quan es desplaça a poca velocitat, el cos adopta una disposició arrodonida, amb la meitat posterior de l’esquena aixecada. Els animals joves tenen un color gris fosc que es perd a la primera muda. Els adults presenten la part dorsal d’un color que varia entre el marró vermellós o el marró grisós i el negrós. Els animals més adults acostumen a presentar tonalitats vermelloses.
La part ventral és blanc brut o grisa, amb els pèls gris clar a la base i blancs a la punta. No hi ha una línia de separació ben marcada entre les coloracions dorsal i ventral.
Biologia
La forma comensal de la rata comuna ocupa els cellers, magatzems i clavegueres. Quan coincideix amb la rata negra, es distribueixen l’espai de manera que aquesta ocupa les parts altes del edificis (fins i tot en un mateix recinte) i la rata comuna habita les parts baixes. La uniformitat de distribució es va perdent a mesura que hom progressa vers els Pirineus. Als llocs més freds, la competència amb la rata negra afavoreix aquesta darrera, que queda com a única resident dels habitacles humans a les zones més elevades. Als Pirineus, la rata negra no constitueix poblacions externes. A les terres baixes, continentals i mediterrànies, la rata comuna genera nuclis poblacionals puntuals que es poden generar temporalment al voltant de les carreteres, en punts on cau gra durant el transport, o d’escombraries; als llocs on hi ha un cultiu permanent d’hortalisses, o d’arròs, també solen instal·lar-se’n poblacions estables. Als Països Catalans, els punts més importants d’aquestes poblacions són les zones de Salses i les ja esmentades del delta de l’Ebre i l’albufera de València, on, fonamentalment, es practica la monocultura de l’arròs, i les zones d’horta de Lleida i Alacant. En aquests indrets, la rata comuna presenta increments poblacionals que provoquen autèntics flagells a l’agricultura. Quan habita en terreny obert, nidifica dins de caus de poca fondària, de 0,5 m aproximadament, que ella mateixa excava als marges de canals i sèquies. De fet, en molts llocs, fa més mal a la infrastructura que als vegetals. Cal considerar-la una espècie omnívora. La seva alimentació varia en funció de les disponibilitats del medi i presenta un ventall de possibilitats molt ampli, tant pel que fa a l’aliment d’origen vegetal com al d’origen animal.
Presenta una activitat primàriament crepuscular. Dins les construccions humanes (cellers, magatzems), hi ha dos màxims curts d’activitat, un després de la posta de sol i un altre abans de la sortida. Els animals joves no segueixen totalment aquest ritme ja que surten tant en cerca d’aliment abans de la posta del sol com a mitjanit i després de la sortida del sol. Els animals que porten vida lliure romanen fora del cau, particularment a l’estiu, fins una hora després de la sortida del sol i, a l’hivern, sobretot amb temps desfavorable, presenten activitat fins i tot al migdia.
A l’Europa central, el 50 % de les femelles assoleixen la capacitat reproductora quan tenen una longitud mitjana de cap i cos de 163 mm i 98 g de pes, i el 50 % dels mascles presenten descens testicular quan fan 159 mm de longitud de cap i cos i 85 g de pes. Aquestes dimensions presenten variacions locals i també en funció de la disponibilitat de l’aliment. En animals criats en captivitat, algunes femelles (poques) crien per primera vegada als tres o quatre mesos. Entre els sis o set mesos, la majoria ja presenta plena capacitat reproductora, que s’atura entre els quinze i els divuit mesos. Durant la seva vida, en captivitat, una femella pot tenir una mitjana de sis a vuit gestacions. Estudis realitzats al delta de l’Ebre, a partir d’animals que viuen fora dels habitacles humans, indiquen que presenten un cicle biològic estacional amb dos màxims d’activitat reproductora, un al maig-juny i un altre al setembre-octubre, més marcat que l’anterior. En els mascles, es detecta la presència d’animals amb capacitat reproductora durant tot l’any. Entre els mesos de febrer i octubre, el 100 % dels exemplars sexualment madurs són actius. Als mesos hivernals, descendeix l’activitat reproductora dels mascles, però n’hi ha un petit percentatge que presenta espermatozoides als testicles.
El cicle d’activitat reproductora comença, en les femelles, al mes de febrer, i al mes de març apareixen ja les primeres gestants. Les darreres femelles gestants apareixen al setembre i a l’octubre encara hi ha alletants, però ja no hi ha indicis d’activitat sexual. Durant els mesos de novembre, desembre i gener cap femella sexualment madura no presenta indicis d’estar sexualment activa. En el cas del delta de l’Ebre, s’observa que la mitjana d’embrions per femella presenta oscil·lacions en funció del mes, de manera que va incrementant a mesura que transcorre l’època reproductora. Al mes de juny, la mitjana és de 8,7 i al mes de setembre de 12,3 embrions. El màxim nombre d’embrions per gestació observat és de setze, però els màxims normals oscil·len entre onze i catorze. Aquest increment progressiu del nombre d’embrions per femella pot estar relacionat amb l’estratègia de supervivència de l’espècie al delta de l’Ebre. La dinàmica intrapoblacional indica que, al llarg de l’any, hi ha unes oscil·lacions importants dels contingents poblacionals. Els mesos hivernals es caracteritzen per una estabilitat que es modifica a la primavera quan s’incorporen els primers joves a la població. Això determina el màxim de primavera. El mes d’agost es caracteritza per una davallada considerable de la densitat poblacional que arriba a límits importants. Aquesta davallada pot ser explicada per la mort d’animals senils que energèticament no sobreviuen a la reproducció primaveral, i per la mortaldat infantil. És realment sorprenent el desmesurat increment poblacional que té lloc al setembre-octubre i que assegura, durant l’hivern, un nivell de contingents poblacionals suficients per a poder portar a terme el següent cicle reproductor. L’increment poblacional del setembre respon a una estratègia reproductora que és afavorida per dos factors interrelacionats: per una banda, el fet, ja esmentat, que al setembre la mitjana d’embrions per femella és molt elevada (12,3) i, per l’altra, el fet que, precisament al setembre, és quan s’ha recollit l’arròs i tot el Delta (carreteres, camps, eres), està ple de grans. Això determina la presència d’una base energètica suficient per a assegurar la supervivència de la gran majoria de les cries que neixen. D’aquesta manera, es recupera ràpidament el contingent poblacional i se supera la gran davallada de l’agost. Això, al Delta (i a indrets comparables com l’albufera de València), es repeteix cada any.
Corologia
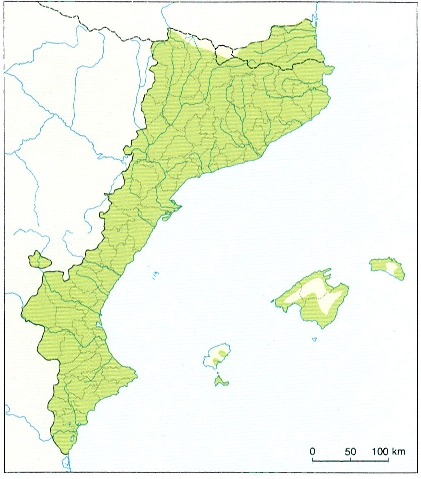
Àrea de distribució de la rata comuna (Rattus norvegicus) als Països Catalans.
Maber, original de l’autor.
La distribució original és paleàrtica, a l’E d’Àsia, on, a les zones temperades de Sibèria i Xina, habiten tant poblacions salvatges com comensals. El camí seguit per la rata comuna en la seva entrada a Europa, probablement durant l’edat mitjana, encara no està totalment aclarit. Les primeres dades precises sobre la seva presència a Europa corresponen al segle XVIII: Anglaterra 1730, França (París) 1753, Dinamarca (Copenhaguen) 1716, Alemanya 1750. A la península Ibèrica, va penetrar a principis del segle XIX. No obstant això, a Europa hi ha restes, trobades en excavacions, que indiquen una arribada anterior. Hi ha restes de ciutats de Polònia dels segles XI-XIII i d’instal·lacions de Scharstorf a Ostholstein posteriors a 1050. Les dades que hi ha sobre la presència de l’espècie al Neolític d’Europa són totalment insegures.
Actualment, la rata comuna colonitza tota Europa. Al N i centre d’Europa, l’ocupació és pràcticament uniforme; en canvi, a l’àrea mediterrània (península Ibèrica, Itàlia i Grècia), la distribució, fora dels nuclis urbans, és dispersa. En aquestes zones, es troba només a llocs particularment humits (Doñana, albufera de València, delta de l’Ebre) o a zones d’horta grans on el regadiu és intens (horta murciana, plana de Lleida). L’aigua és un requeriment molt important de l’espècie i arriba a condicionar la seva presència. A l’albufera de València ocupa la zona humida, pròpiament d’albufera, i la rata negra ha estat desplaçada i ha passat a ocupar la zona de bosc que, sobre un terreny més sec i sorrenc i amb vegetació pròpiament mediterrània, hi ha entre l’albufera i el mar. Contra el pronòstic d’alguns autors de començament de segle, les zones d’ocupació de la rata comuna no han experimentat un increment sensible en els darrers anys.
Als Països Catalans, la rata comuna viu pràcticament arreu on hi ha un habitatge humà. La presència de l’espècie, però, no és regular. Hi ha dos tipus de factors que la condicionen i que afecten independentment les poblacions estrictament comensals i les que viuen al camp, fora dels habitacles humans: la competència amb la rata negra i els factors ambientals, per una banda, i la presència d’aigua, per l’altra. Cal potser finalitzar el tractament d’aquesta espècie recordant que la rata blanca, tan utilitzada en experimentació de laboratori, és la varietat albina de Rattus norvegicus.