Característiques del grup
L’ordre dels lepidòpters el formen les anomenades papallones. El fet de tractar-se dels insectes més vistents ha contribuït a fer-los força populars i a ésser coneguts i col·leccionats per molta gent, i fins a ésser criats (cucs de seda); però les arnes que es menjen la roba, la farina i la cera, el cuc de la col, el del blat, el barrinador de l’arròs, la tinya de la patata, el corc de les pomes i, també, la processionària que infesta els nostres boscos de pins, també són lepidòpters. Veiem doncs, que l’àmbit d’aquest ordre és gran i, per tant, complex, i això fa que en aquesta obra només en puguem tractar els aspectes més generals. De fet, constitueixen un grup força homogeni pel que fa als caràcters fonamentals, caràcters que s’han mantingut al llarg de milions d’anys d’evolució (els fòssils trobats del Secundari ja es poden classificar dins de les famílies actuals); aquesta constància estructural i funcional no ha impedit que s’originessin gran quantitat de formes gràcies a l’adaptació de les espècies als diferents ambients.
Els noms vulgars que hom ha atribuït als diferents representants d’aquest ordre són múltiples, en concordança amb la seva variabilitat de formes, però només citarem els aplicats als grans grups de lepidòpters. A part els ja indicats, hom pot destacar els noms de paloma, voliaïna, pampallola i d’altres derivats de papallona, falena (que s’aplica a les nocturnes), borinot (mot que també s’usa per a designar alguns himenòpters), esfinx i bufaforats (que es refereix a diversos esfíngids), pedraferits (que són les erugues glauques més vistents), i blavetes (que hom aplica als mascles d’alguns licènids i, per extensió, a tota la família).
Després dels coleòpters, l’ordre dels lepidòpters és el més nombrós dels insectes i, àdhuc, dels éssers vius. Això és un aspecte de l’enorme variabilitat a què ens referíem abans. La uniformitat estructural, però, permet distingir fàcilment les espècies d’aquest ordre de la resta d’insectes. L’ordre més semblant és el dels tricòpters, dels quals es diferencien perquè aquests són recoberts essencialment de pèls, més que d’escates. El nombre d’espècies de lepidòpters conegudes supera els 150 000, però, donat que constantment se’n descriuen de noves i que hi ha grans zones del món poc o gens estudiades, hom pot assegurar que el seu nombre real ultrapassa les 200 000 espècies; d’aquestes, a Europa en viuen unes 8000. Pel que fa als Països Catalans, en un càlcul aproximat, situaríem el nombre d’espècies entre 4500 i 5000, de les quals, però, únicament en podriem catalogar unes 2000, atès que els microlepidòpters, que en constitueixen la major part, han estat poc estudiats al nostre país.
Des del punt de vista morfològic, la característica més notable dels adults, i que dona nom al grup, és que tenen les ales i altres parts del cos recobertes d’escates (pteron, ala, lepido, escata). A més, tenen quatre ales membranoses, grosses en relació a les dimensions del cos (envergadura alar d’1 mm a 30 cm) i proveïdes de nervacions. Les peces bucals formen l’anomenada espiritrompa. Les femelles ponen, mitjançant un orifici diferent del copulador, nombrosos ous molt petits i de formes molt diverses, dels quals emergeix una larva eruciforme (l’anomenada eruga) proveïda de mandíbules i de cinc parells de falses potes a l’abdomen, a més dels tres parells de potes toràciques articulades; la pupa, que s’anomena crisàlide, és compacta, gairebé immòbil, i sovint és voltada d’un capoll elaborat per l’eruga amb fils de seda que segrega ella mateixa.
Des del punt de vista ecològic, els lepidòpters són productors secundaris d’organització complicada, micròfags de superfície, i formen part del segon nivell tròfic dels ecosistemes terrestres. La seva importància ecològica és notable, bàsicament perquè s’alimenten de plantes i tenen una taxa de reproducció molt elevada, per la qual cosa n’hi ha que provoquen flagells a l’agricultura, d’altres que defolien els boscos, si bé sovint és a causa de desequilibris ocasionats per l’home. Tanmateix, no podem deixar d’assenyalar, en sentit contrari, l’eruga de Bombyx mori, el cuc de seda, que es cultiva, a la Xina, des de fa més de 4000 anys.
Morfologia de l’adult
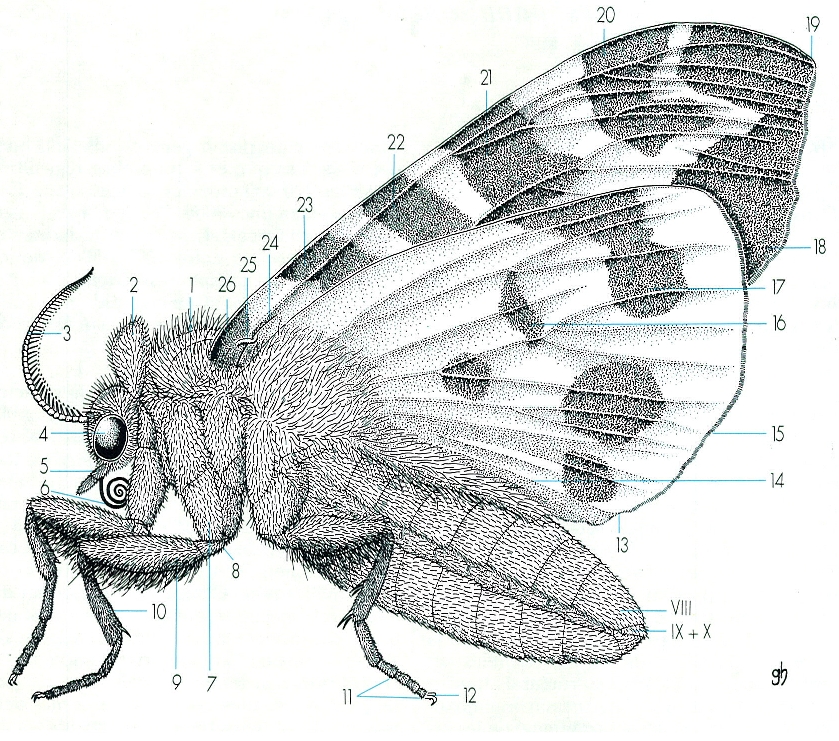
Aspecte general extern d’un lepidòpter (×8), en visió lateral, en el cas d’un àrctid (Arctia caja): 1 tòrax, 2 collar, que separa la càpsula cefàlica del tòrax, 3 antena, 4 ull compost, 5 palp labial, 6 espiritrompa, 7 coxa, 8 trocànter, 9 fèmur, 10 tíbia, amb un esperó tibial, 11 tarsos, 12 ungles, 13 angle anal, 14 marge anal, 15 fímbries, 16 taca reniforme, 17 banda antemarginal, 18 banda marginal, 19 àpex, 20 banda extramediana, 21 banda mediana, 22 banda antemediana, 23 banda extrabasilar, 24 fre, 25 retinacle, 26 zona basal.
Gustavo Hormiga, del natural.
Des del punt de vista estructural, la morfologia dels imagos de lepidòpter és la típica dels insectes evolucionats. Hom en descriurà cada part del cos, tot indicant els apèndixs i els òrgans sensorials que s’hi relacionen. Potser una de les característiques més destacables dels lepidòpters és la presència d’escates, que, si bé n’hi ha per tot el cos, és a les ales on resulten més evidents, imbricades d’una manera semblant a les teules d’una teulada.
Les parts del cos
El cap dels lepidòpters és una petita hemisfera amb poc moviment, que duu la boca i, a més, òrgans sensorials tan importants com els ulls compostos, capaços de captar radiacions ultraviolades, i les antenes; molts heteròcers també tenen dos ocel·les. Les antenes són apèndixs allargats, compostos d’artells, i de formes molt diverses. Una classificació dels lepidòpters molt utilitzada, els separa en ropalòcers (amb antenes filiformes acabades en maça) i heteròcers (antenes d’altres formes). El sentit preponderant que poseeixen les antenes és l’olfacte, però també tenen tacte. En les espècies que basen l’atracció sexual en l’olor, són les antenes del mascle les encarregades de captar les ínfimes quantitats de feromona que la femella emet.
Les peces bucals s’han modificat extraordinàriament, i conformen un dels òrgans més típics de l’ordre: l’espiritrompa. En posició de repòs, l’espiritrompa roman plegada, gràcies a la seva pròpia elasticitat, com la corda d’un rellotge, i quan l’insecte vol xuclar un suc nutritiu, es desenrotlla fins a atènyer una longitud superior a la total de l’imago (en alguns esfíngids, com Agrius convolvuli, arriba a ser el doble); això els permet arribar als nectaris més profunds de les flors, fora de l’abast d’altres insectes. Així, els lepidòpters actuen de pol·linitzadors de les plantes fins al punt de crear una relació d’interdependència que ha arribat a la coevolució, de la qual parlarem més endavant. L’extrem de l’espiritrompa és proveït d’òrgans sensorials i, en algunes espècies tropicals, és rígid i dentat per tal de foradar els teixits vegetals. Hi ha papallones que no tenen espiritrompa o la tenen més o menys atrofiada, de manera que quan són adults no es poden alimentar.
A ambdós costats de l’espiritrompa hi ha els palps, en alguns casos molt evidents (per exemple, els cràmbids i alguns noctúids), de funció sensorial (tacte i/o olfacte). El sentit de l’olfacte també el proporcionen els òrgans quimioceptors, inclosos en petites cavitats situades als palps. El gust el capten quimioceptors presents a l’extrem de l’espiritrompa i, de vegades, a les potes; és conegut l’experiment de mullar les potes d’un nimfàlid o pièrid amb una solució ensucrada, al qual el lepidòpter respon desenrotllant l’espiritrompa i cercant la situació de l’aliment. La visió es realitza per mitjà dels ulls compostos i els ocel·les, però no tenen particularitats diferenciadores de la resta dels insectes. Direm, tan sols, que capten radiacions ultraviolades. El tacte prové de les setes tàctils, que són uns pèls sensitius repartits per tot el cos, però concentrats en els palps, les antenes i les potes, que detecten el tipus de superfície, i arriben a distingir una branca d’una fulla, per exemple. L’audició es concreta de les maneres més diverses; els grups ancestrals tenen sistemes molt simples, però a mesura que es puja en l’escala evolutiva, anem trobant òrgans cada vegada més complexes: els nimfàlids ja tenen una membrana sensible, els esfíngids disposen d’un òrgan acústic als palps, geomètrids i piràlids mostren un autèntic timpà situat a la part anterior de l’abdomen, i els noctúids han desenvolupat un òrgan timpànic al metatòrax. En els casos més perfeccionats poden captar ultrasons de més de 80 000 Hz, cosa que els permet fugir dels rats-penats, que usen l’ecolocació amb ultrasons.
El tòrax, com a suport dels òrgans de locomoció, és la part més rígida del cos, i consta de tres segments: el protòrax, que suporta el primer parell de potes (les més febles); el mesotòrax, on s’articula el segon parell de potes i el primer parell d’ales (les més grosses); i el metatòrax, portador del tercer parell de potes i el segon parell d’ales, i, en algun cas (com els noctúids), d’un parell d’òrgans timpànics.
Diferentment del que passa a la majoria d’insectes, en els lepidòpters les potes no solen servir per a caminar: no són robustes, i llur funció és mantenir l’equilibri en repòs; només ocasionalment les fan servir per a córrer curtes distàncies, i també són útils per a conservar l’estabilitat en l’aterratje. Poden ésser cobertes d’escates i pèls, amb major o menor densitat. Hi ha espècies que hi tenen l’anomenada epífisi tibial, un esperó de la tíbia amb el qual es netegen les antenes. Alguns ropalòcers (com, per exemple, els nimfàlids) són tetràpodes per atròfia del primer parell de potes, i un cas extrem és el de les femelles d’alguns psíquids, que són totalment àpodes.
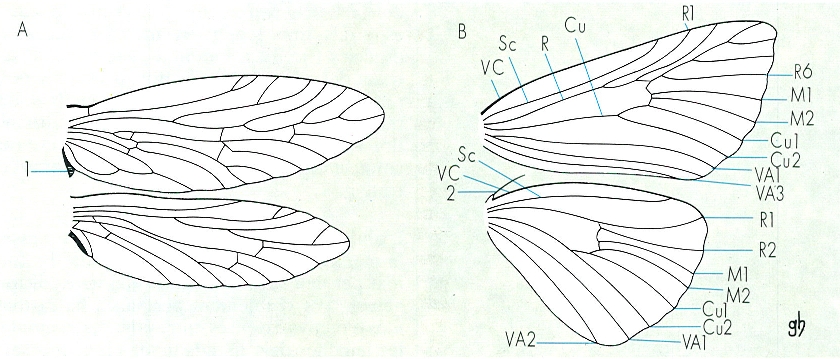
Estructura de les ales i nervació alar en els lepidòpters, segons la qual hom distingeix els homoneures(A) dels heteroneures (B). Els homoneures o isoneures tenen un sol patró de nervació, que és semblant al d’altres ordres d’insectes, per tal com són els lepidòpters menys evolucionats: també s’anomenen jugats (Jugatae) perquè acoblen les ales per mitjà del jou (1). Els heteroneures o anisoneures, de nervació modificada i diversa, solen tenir fre (2), per la qual cosa també reben el nom de frenats (Frenatae); són els lepidòpters típics. Hom consigna en el dibuix: VC vena costal, Sc subcostal, R radial, Cu cubital, M mediana, VA anal.
Gustavo Hormiga, a partir de fonts diverses
Les ales dels lepidòpters són molt visibles i característiques de l’ordre i no només permeten un vol força actiu, sinó que, a més, representen un sistema idoni per a difondre calor en cas de temperatures altes i per a captarne en llocs assolellats quan fa fred. La distribució dels nervis alars segueix determinats models que els especialistes empren en la taxonomia, raó per la qual cada nervi té un nom, així com cada zona que delimita. Les famílies més primitives (micropterígids i hepiàlids) són homoneures, és a dir, tenen la nervadura de les ales anteriors igual que la de les posteriors, mentre que la resta de lepidòpters són heteroneures. Per tal que es moguin sincrònicament, les ales de cada costat del cos estan acoblades per sistemes especials, que tenen importància taxonòmica. Els homoneures tenen una llengüeta, anomenada jou, a l’ala anterior, que encavalca sobre l’ala posterior. Els heteroneures habitualment disposen del fre: un pèl gruixut (més d’un i més prims en les femelles) que surt de l’ala posterior i s’acobla a un conjunt de pèls curts que hi ha a l’ala anterior. Els ropalòcers (i alguna família d’heteròcers) tenen el sistema anomenat amplexiforme, que consisteix en una dilatació de la base de l’ala posterior, que s’aplica sobre l’anterior. La forma de les ales és molt variable, i obeeix, en cada cas, a diferents adaptacions.
La distribució de les escates sobre les ales determina diferents dissenys de colors, i la infinitat de dibuixos possibles fa que hom pugui considerar-hi tota una tipologia. Indicarem alguns dels noms més usats en aquest camp: traç (línia llarga), banda o franja (línia ample), estria (línia curta i fina), ombra (coloració difuminada), ocel·le (dos o més cercles concèntrics), lúnula (forma de mitja lluna), malgrat que sempre es donen situacions intermèdies que cal especificar en cada cas. Els dissenys alars serveixen per a la comunicació entre individus: per a la identificació de sexes i espècies, per al camuflatge, per a advertir de la seva toxicitat, per a espantar els depredadors, etc.; a més a més, un mateix individu pot comunicar diferents missatges movent les ales, plegant-les de diferent manera, etc.
Estructuralment, l’escata és formada per dues làmines unides per trabècules, amb un gruix total de l’ordre de mitja mil·lèsima de millímetre. D’escates n’hi ha de moltes formes i colors. Habitualment, el color el proporcionen els pigments cuticulars de la mateixa escata (melanines, pterines, etc.), però també pot ésser causat per un efecte òptic, produït per difracció, reflexió, o per interferència de la llum en travessar la superfície irregular d’una escata especial, que precisament sol ésser despigmentada; la interferència prové del fet que les arestes de les estries de l’escata són separades entre elles per distàncies inferiors a la longitud d’ona de la llum, mentre que la difracció l’ocasionen les finíssimes làmines de l’escata separades per capes d’aire, que fan que el raig de llum es trobi amb índexs de refracció alternativament diferents. A més, en variar la inclinació de l’eix de la llum, aquesta es descompon en els diferents colors, i així s’originen les típiques tonalitats metàl·liques, blaves, verdes, tornassolades, violàcies, iridiscents, etc., que varien amb els moviments de l’ala. Els androconis són grups d’escates especials que tenen els mascles d’algunes espècies (licènids, nimfàlids, etc.) a diverses parts del cos, però, sobretot, a les ales posteriors; aquestes escates androconials solen tenir forma de plomall per tal de difondre millor l’olor que prové d’una glàndula interna connectada amb l’escata i que actua durant el festeig.
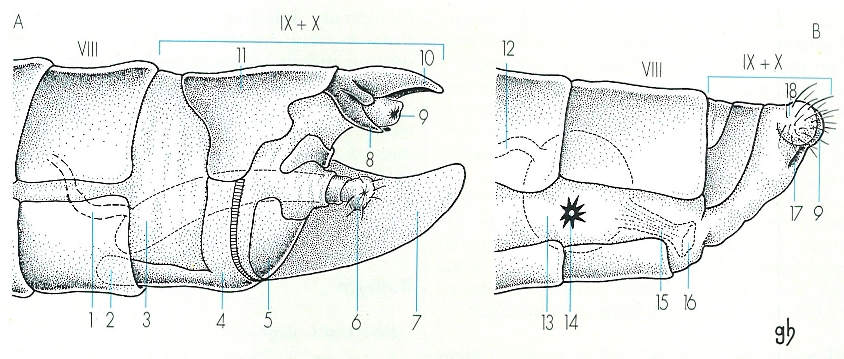
Extrem de l’abdomen i genitàlia dels lepidòpters. Dels tres tipus fonamentals presents en els lepidòpters, és a dir, el monotrisià, (amb un únic orifici cloacal), l’exoporià i el ditrisià, aquí representem aquest darrer, el més típic i evolucionat, en el cas d’un mascle (A) i d’una femella (B). Les característiques de la genitàlia masculina o andropigi i de la femenina o ginopigi són emprades en la taxonomia. Hom consigna els elements morfològics següents: 1 canal ejaculador, 2 sacus, 3 edeagus, 4 vínculum, 5 juxta, 6 vesica, 7 valva, 8 gnatos, 9 anus, 10 uncus, 11 tegumen 12 conducte seminal, 13 bossa copuladora, 14 signe, 15 conducte de la bossa copuladora, 16 orifici copulador, 17 ovipositor, 18 papil·les anals.
Gustavo Hormiga, a partir de fonts diverses
L’abdomen, la darrera part del cos, sol ésser la més grossa i la menys quitinitzada. És format per deu segments, però mai no es distingeixen tots visiblement perquè hi ha fusions més o menys extenses entre ells; per exemple, les femelles dels còssids tenen els últims segments modificats en forma de barrina formant l’ovopositor; en els geomètrids, el primer segment porta un parell d’òrgans timpànics. A l’abdomen es localitza la majoria de funcions fisiològiques de l’individu, i al seu extrem hi ha l’aparell reproductor, la part externa del qual és esclerificada i forma l’anomenada genitàlia. Als mascles, hom hi pot distingir les dues valves que, a l’acoblament, subjecten l’abdomen de la femella. Les femelles tenen l’abdomen acabat en punta, i el més notable del seu aparell reproductor és que les funcions de còpula i de posta es fan per orificis diferents (situats respectivament al vuitè i al novè segment); són excepció a aquesta característica les papallones més primitives, que tenen un sol orifici, pel qual fan ambdues funcions (monotrisians).
El polimorfisme

El dimorfisme sexual en els macrolepidòpters adults sovint és molt evident, com en Lasiocampa quercus, espècie que recull la fotografia: el cos i les ales del mascle solen ésser més petits i, de vegades, a més del color, també varia el disseny alar. Les femelles tenen l’abdomen més gruixut i l’extrem més agut, de vegades amb l’ovipositor llarg, i sense la tofa de pèls llargs. Els lepidòpters amb les ales reduïdes o absents (àpodes) sempre són femelles. Si presenten escates androconials, en canvi, sempre són mascles. En l’estat de crisàlide en alguns casos (satúrnids, noctúids, etc.) ja es pot endevinar el sexe per la morfologia dels darrers segments de l’abdomen o pel relleu de les antenes (més pectinades en els mascles).
Jordi Vidal/Col·lecció Vilarrúbia.
Sense esser-hi tan desenvolupat com en altres insectes (pensem en les abelles, les formigues, els tèrmits, per exemple), en els lepidòpters també hi ha un cert grau de polimorfisme. Hom en distingeix dos tipus: el sexual i l’estacional.
El polimorfisme sexual es concreta habitualment en el típic dimorfisme sexual, tan generalitzat a la majoria dels grups d’animals, pel qual els caràcters sexuals secundaris són els més evidents, i es manifesten sobretot en els imagos, cosa lògica per tal com és la fase reproductora. En els lepidòpters, en general, es reconeixen fàcilment els individus d’un i altre sexe i en alguns grups s’observa un gradient de diferenciació: en els geomètrids hi ha femelles braquípteres (amb les ales atrofiades i no funcionals), micròpteres (ales reduïdes a simples munyons) i àpteres (desaparició absoluta de les ales). El cas extrem de dimorfisme el trobem en el psíquid Psyche unicolor, la femella del qual és àptera i àpoda, d’aspecte més semblant a un cuc, que no a una papallona. Entre els lepidòpters del nostre país tenim un cas d’autèntic plurimorfisme sexual: es tracta del pièrid Colias crocea, del qual hom coneix quatre formes: una única del mascle i tres de femelles (la típica, l’helice i l’helicina), a més de formes intermèdies.
El polimorfisme estacional es dona en algunes espècies plurivoltines i, sense ésser un fenomen gaire estès ni gaire acusat, és freqüent a determinades famílies, com els pièrids i els licènids. No s’ha de confondre amb les diferències que s’observen en alguns imagos hivernants, com Nymphalis antiopa, els quals, després d’haver passat l’hivern, tenen tonalitats menys vives que les dels individus acabats de néixer. El que sí que és un cas d’autèntic polimorfisme estacional, i força acusat, és el del nimfàlid Araschnia levana, present al N dels Països Catalans, la generació d’estiu del qual té dissenys i colors totalment diferents dels de la generació de primavera, que els ha engendrat; la diferència és tant notable, que sempre s’havia cregut que eren dues espècies diferents, fins que els cultius demostraren que les unes donaven lloc a les altres.
Finalment, indicarem un tipus de polimorfisme especial, restringit a l’etapa larval, en què una mateixa espècie produeix erugues diferents. Per exemple, de l’esfíngid Agrius convolvuli podem trobar erugues negres, marrons i verdes, i no a causa de l’alimentació, com passa en altres espècies. Aquesta diversitat polimòrfica, que, de fet, és policromia, no es reflecteix en els adults, els quals són tots iguals.
Biologia
Els cicles biològics
Als països tropicals és corrent que les fases del cicle biològic dels lepidòpters es vagin succeint al llarg de l’any sense parar, però, a les nostres latituds, en què el clima sofreix canvis importants (estacions), els lepidòpters, més adaptats al temps calorós, es veuen obligats a interrompre el desenvolupament durant l’hivern. Qualsevol de les quatre fases pot ser l’encarregada de realitzar la diapausa, encara que les més idònies, per llur protecció i manca de necessitats, són la crisàlide i l’ou. Això i la necessitat de realitzar la desclosa de l’ou quan la planta hoste ja ha brotat, i el naixement de l’adult quan la planta nutricia ha florit, condiciona l’existència d’un ritme anual. Per a saber en quin moment de l’any es troben, els lepidòpters capten el fotoperíode, bé que també es guien per altres factors, com la temperatura i la pressió atmosfèrica. Per a conèixer el fotoperíode, algunes espècies han desenvolupat sistemes sorprenents, com el d’Antheraea pernyi, un satúrnid exòtic introduït a Mallorca, on encara es troba esporàdicament, la crisàlide del qual té una finestra transparent que capta i mesura la llum. A determinades espècies (com la raça bivoltina de Bombyx mori), els individus saben si han de fer diapausa o no gràcies a la informació presa per la generació anterior, que és la que ha estat exposada a llargues il·luminacions i ha sintetitzat la substància desencadenant.
Les espècies univoltines, que són la majoria, són les que completen el cicle cada any; les plurivoltines el completen més d’una vegada l’any (bivoltines si hi ha dues generacions, trivoltines si n’hi ha tres, etc.). La durada del cicle és molt variable: des de poques setmanes, cas en el qual les espècies, de cicle ràpid, poden tenir moltes generacions cada any (en un nombre no fix), fins a alguns anys, com passa a Cossus cossus. La papallona de la col (Pieris brassicae) pot tenir més generacions de les usuals si s’allarga el bon temps.
El control de totes les fases de creixement al llarg del cicle biològic és hormonal, i és d’una gran complexitat. Hi ha hormones encarregades d’anar determinant periòdicament les mudes, d’altres que determinen el moment de crisalidació, etc.
La reproducció
Entre les pautes de conducta de les papallones en relació amb la reproducció, poden separar-se les específiques dels mascles, les de les femelles, i les conjuntes, encaminades al trobament de la parella.
Del comportament típic dels mascles destacarem la dansa nupcial i la territorialitat. La dansa nupcial es coneix especialment en els mascles dels ropalòcers, als quals no opera l’atracció per feromones femenines; però en qualsevol cas, mai no és tan complexa com la dels ocells, ni tan coneguda. Pel que fa a la territorialitat, es coneix especialment a les espècies diürnes; els mascles solen prendre el sol a les clarianes del bosc, i quan un altre individu entra al territori, si és femella, poden iniciar la dansa nupcial, però si és un altre mascle, inicien tot un ritual per a foragitar-lo.
La recerca de la parella en les espècies diürnes és basada en l’atracció visual dels colors i els dissenys alars. Inicialment, els mascles poden seguir qualsevol espècie i fins i tot objectes de colors; és típica l’escena d’un mascle seguint fins a terra una fulla que cau de l’arbre. La identificació es produeix després de l’apropament i, si és positiva, comença el vol nupcial. L’eficiència és, doncs, a curta distància.
Les espècies nocturnes han desenvolupat altres sistemes d’atracció, dels quals destaquen les feromones sexuals. Des de fa segles, els entomòlegs havien observat la misteriosa atracció que les papallones femelles exercien sobre els mascles, i havent descartat el concurs de la vista i l’oïda, prodigaven explicacions fantàstiques, fins fa ben poc (1960), que s’aïllà la primera feromona que confirmà la hipòtesi de les substàncies oloroses específiques emeses per les femelles. Actualment, però, hom és conscient que el mecanisme és més complicat, i que hi intervenen, almenys, dues molècules, una excitadora i l’altra inhibidora. L’estructura química de les feromones no és molt complexa, sinó que normalment es tracta d’hidrocarburs de 10 a 18 àtoms de carboni, amb algun grup funcional; si les molècules fossin més petites, no permetrien la variabilitat necessària per a l’especificitat de cada espècie; més grosses, perdrien volatibilitat i, en conseqüència, poder d’atracció. L’extraordinària eficiència del sistema no solament és conseqüència de la facilitat de dispersió de les feromones a llarga distància, sinó també de la gran sensibilitat per a captar-les que tenen els mascles. Una sola molècula que toqui els receptors de les antenes, ja fa enviar un impuls nerviós; tanmateix, per a superar la "remor de fons" que té qualsevol òrgan sensitiu, es necessiten uns quants impulsos i la quantitat mínima de feromona per a que el mascle capti la seva presència es calcula al voltant d’1 mg per 100 km3. Però, per a poder-se orientar, els mascles han de localitzar el gradient de concentració fins a trobar la femella, la qual pot trobar-se fins a alguns quilòmetres de distància. La persistència de l’efecte també és notable: un mascle del limàntrid Orgyia antiqua pot acudir i intentar l’acoblament amb una femella que fa uns quants dies que és morta. Les femelles no sempre emeten feromones, i la quantitat amb què ho fan depèn de l’edat, del temps atmosfèric, de l’hora i del seu grau de fecundabilitat. Els avantatges del sistema d’atracció per feromones són múltiples: perdura més que els senyals visuals i acústics, té un cost energètic menor i evita la localització per part dels depredadors, que no en poden seguir el rastre. A més, el fet que sigui el mascle el qui acudeixi, permet que la femella es dediqui plenament a cercar la planta nutricia per a dipositar-hi els ous, factor que és essencial per a la perduració de l’espècie. Malgrat això, ja hem indicat el cas dels mascles d’alguns nimfàlids que emeten feromones, però que les usen, més que per atreure la femella, per a fer-la més receptiva durant la dansa nupcial. Això no obstant, hom interpreta com a poder d’atracció per a les femelles l’olor dels mascles d’alguns hepiàlids, i la remor que produeixen determinats noctúids en reunir-se els mascles i picar-se les ales entre ells.
Abans d’iniciar l’acoblament, algunes femelles necessiten alimentar-se per a aconseguir la maduresa sexual; d’altres també necessiten passar una diapausa hivernal o estival. Les que neixen amb els òvuls madurs no solen alimentar-se gens, i algunes copulen i fan la posta immediatament després del naixement, estratègia que eludeix el risc que els depredadors impedeixin la posta. La durada de la còpula pot ser de pocs minuts fins a més de dos dies, i pot ésser única o múltiple, immòbil o en mig de grans volades, a vegades sorprenentment hàbils.
Els mascles solen morir poc després de l’acoblament (o acoblaments amb diverses femelles), però els que no aconsegueixen aparellar-se tampoc no viuen gaire més. Les femelles moren poc després de la posta. Una excepció la constitueixen els imagos que hivernen.
La posta
Després de l’acoblament, la funció més important de la femella és realitzar la posta. Les espècies polífagues solen deixar caure els ous en ple vol, mentre que les monòfagues o oligòfagues solen tenir més cura en seleccionar la planta, i de vegades deixen els ous un per un en fulles diferents, com el cas del nimfàlid Charaxes jasius. El més normal, però, és que els ponguin agrupats, enganxats amb un vernís especial, sobre la planta hoste. El nombre d’ous acostuma a ser d’alguns centenars, bé que hi ha casos de més d’un miler, i fins de 4000 en el noctúid Spodoptera littoralis, si bé una part dels ous de la posta sol ésser estèril (menys del 5 %). Si la femella no aconsegueix d’acoblar-se, també efectua la posta, en aquest cas, estèril en un 100 %; tanmateix, això té una excepció en alguns psíquids, que presenten partenogènesi.
L’ou és la primera fase embrional, i sol durar poc. Encara que hi ha casos en què hiverna i es paralitza així la formació de l’eruga, la durada normal és d’uns quants dies. En algunes espècies, l’eruga eclosiona un parell d’hores després de la posta i, en conseqüència, neix petitíssima. Hom pot preveure el moment de l’eclosió perquè la coberta és transparent. La forma, i àdhuc els colors, dels ous de lepidòpter és molt variable; habitualment, el còrion té relleus i ornamentacions, fins al punt que, sovint, hom pot classificar l’espècie només observant-ne l’ou, a gran augment.
L’eruga i la crisalidació
La finalitat principal de l’eruga és alimentar-se i acumular reserves per a les transformacions i la vida posteriors. La seva morfologia i el seu comportament són encaminats a assolir aquest objectiu. Estructuralment, les larves de lepidòpter no difereixen gaire de les dels altres insectes, però la seva forma externa, força característica, ha fornit l’adjectiu "eruciforme", que hom aplica a totes les formes semblants.
La gran variabilitat de l’ordre dels lepidòpters es reflecteix d’una manera molt clara en la immensa diversitat de formes d’erugues. N’hi ha de glabres, amb pubescència uniforme, acumulada en pinzells, modificada en espines, amb pèls urticants, etc.; n’hi ha que tenen protuberàncies més grosses que la llargada total del cos. Quant a la pigmentació, no cal dir que tots els colors hi són presents: des dels tons apagats que faciliten el camuflament, fins als més vius aposemàtics (no són rars els casos de toxicitat), i amb els dibuixos més variats. Sovint, la coloració és provocada o modificada per l’alimentació: en les espècies polífagues, les erugues d’una mateixa posta poden adquirir diferents colors si mengen plantes diferents.
Després del cap, que és ben diferenciat, els tres segments del tòrax es distingeixen fàcilment pel parell de potes articulades que porta cadascun. A continuació, l’abdomen consta de deu segments, dels quals els dos o tres darrers solen ésser fusionats, i de vegades modificats en forma de ventosa, de cua enlairada, de protuberàncies, etc. Normalment, hi ha parells de falses potes o pseudopodis situats a cinc segments abdominals (poden tenir-ne més o menys; els geomètrids només en tenen dos parells). Les falses potes són expansions membranoses acabades en ventosa, i la seva finalitat és adherir fortament l’eruga al substrat. A cada costat del gran tub digestiu hi desemboquen les dues glàndules sericígenes que fabriquen la seda i l’emmagatzemen en estat líquid. Aquesta seda, en prendre contacte amb l’aire, i amb l’ajut d’altres glàndules accessòries, se solidifica, tot conservant la flexibilitat, i forma el fil de seda, que consta de dos brins de fibroïna, envoltats per sericina i altres substàncies; el resultat és d’una resistència sorprenent (50 kgr/mm2) comparable a la de l’acer. L’eruga utilitza la seda per a fixar-se a la planta nutricia, per a despenjar-se amb seguretat en canviar de mata i per a construir el capoll que protegirà la crisàlide. Pel que fa als òrgans sensorials, són força reduïts. Els ulls simples només permeten captar intensitats lluminoses. La cutícula porta pèls i vesícules que proporcionen a l’eruga els sentits de tacte, olfacte i oïda.
La voracitat de l’eruga és notable, i també el seu creixement (pot incrementar de pes unes deu mil vegades). El creixement no és continu, sinó dividit, habitualment, en quatre o cinc fases, separades per les mudes; hi ha casos de més de deu mudes, així com erugues de microlepidòpters (gènere Tineola) que muden sense crèixer. L’aspecte de la larva pot anar canviant, i hom pot endevinar la proximitat d’una muda en veure la nova pell (amb dissenys i colors eventualment diferents) per transparència sota l’exúvia vella. Llevat de les hivernants, la durada normal de la fase d’eruga és d’unes quantes setmanes; algunes, però, amb pocs dies de vida en tenen prou, i d’altres no crisaliden fins al cap d’uns anys. Aquest és el cas de Cossus cossus; atès el fet que es tracta d’un lepidòpter gros, l’eruga neix amb una mida molt reduïda, i la fusta de què s’alimenta és poc nutritiva.
Les erugues es mouen aixecant el darrer segment i encorbant el cos. Aquesta curvatura es va traslladant cap endavant, elevant tots els segments del cos fins arribar al cap. Aquest és el procediment normal, però hi ha variacions; per exemple, els geomètrids se salten els passos intermedis i col·loquen l’últim segment directament al costat del cap.
La crisalidació és el pas de la darrera fase d’eruga a l’estat nimfal tot realitzant l’última muda. Igual que en les altres mudes, l’eruga deixa de menjar, però ara elimina tots els excrements i perd d’un 20 a un 50 % del seu pes; després de buscar un lloc arrecerat o d’enterrar-se, elabora l’embolcall pupal i es transforma en crisàlide. Els ropalòcers i altres grups no construeixen capoll ni s’enterren sinó que es penjen, per exemple, d’una fulla o una branca; tanmateix, alguns hespèrids i satirins també construeixen embolcall.
La crisàlide
La fase nimfal dels lepidòpters s’anomena crisàlide, i per tal com és un estadi de profunds canvis interiors, però de poc moviment extern, requereix un sistema de protecció. El més habitual és l’elaboració d’un embolcall pupal, que pot fer-se a sota terra o a llocs més o menys amagats (entre l’escorça dels arbres, a les escletxes de les roques, etc.), de vegades construït de seda reforçada amb fragments vegetals (fulles seques, branquillons, etc.), i d’altres (en famílies que s’enterren) que, en comptes de fer capoll, construeixen un embolcall de fang amb saliva. Un altre tipus de crisàlide és la dels ropalòcers, els quals usen la seda només per a penjar-se de l’extrem de l’abdomen i, eventualment, fent una mena de cinturó fixat al substrat. Donat que no hi ha capoll, solen situar-se a llocs arrecerats i són de colors apagats i de camuflament. La longitud del fil de seda secretat per l’eruga pot arribar a ésser d’alguns quilòmetres i d’un diàmetre d’una mil·lèsima de mil·límetre.
Quant al moviment de la crisàlide, és molt restringit, i es limita normalment a rotacions abdominals. Examinant la superfície de la crisàlide, hom pot distingir, pel relleu format a la cutícula, els òrgans que tindrà l’adult (els ulls, l’espiritrompa, les ales plegades, les antenes, etc.), i, a diferència del que passava a l’eruga, en molts casos s’arriba a discernir el sexe de l’exemplar per diferents detalls: l’empremta de les antenes, més grossa en els mascles; les dimensions generals de la crisàlide (les femelles solen ésser més grosses), els dos orificis genitals (als segments vuitè i novè) de les femelles i l’orifici únic (al segment novè) dels mascles. Tot el que hem dit és vàlid pel tipus de nimfa adèctica obtectada, al qual correspon la immensa majoria de lepidòpters. Però cal assenyalar el cas de les famílies més primitives (micropterígids, eriocrànids i, més restringidament, els hepiàlids), que tenen nimfa dèctica o lliure, amb les antenes desenvolupades i no adherides al cos, i les mandíbules mòbils (fins i tot són funcionals en els micropterígids).
La grandària de les crisàlides oscil·la entre els 2 mm (alguns microlepidòpters minúsculs) fins als 70 mm (els esfíngids més grossos). Quant a la durada de l’estat pupal, el més normal és que sigui de poques setmanes, encara que hi ha casos en què és d’alguns dies, i no és rar que hiverni; aleshores dura uns quants mesos; també es troben espècies en què dura anys. La durada pot variar dins la mateixa espècie i, de fet, canvia molt sovint, segurament perquè és una estratègia més adequada per a salvar situacions ambientals desfavorables. En espècies en què sol durar pocs mesos, s’han detectat casos en què l’estat nimfal s’ha perllongat fins a 6 anys i, després, l’adult ha nascut perfectament.
El naixement de l’imago
Excepte en els casos de prolongació més enllà de la temporada, el desencadenament dels processos que donen lloc a finalitzar la formació de l’insecte adult ve determinat per les variacions climàtiques de l’indret i pel fotoperíode. Per això, en un lloc concret, la majoria d’imagos solen néixer durant un període de pocs dies. Ara bé, una vegada format l’adult, el moment exacte de la desclosa de la crisàlide no es produirà fins que les condicions microclimàtiques de l’ambient siguin idònies. Per a evitar néixer en un dia de pluja, la crisàlide detecta la temperatura i la pressió atmosfèrica, que indiquen el bon temps; per aquesta raó, en dies calorosos de primavera és normal notar, de sobte, una superpoblació d’imagos de la mateixa espècie. En el primer dia de bon temps que segueix uns dies ennuvolats és més fàcil que es donin aquestes sortides massives. Aquesta simultaneïtat pot arribar a fer coincidir l’emergència dels adults en una hora del dia que, en els ropalòcers, sol ser a primera hora del matí.
Una de les primeres coses que fa l’adult en néixer és expulsar el meconi, que és el líquid que conté els materials sobrers produïts per la crisàlide i que aquesta no ha pogut eliminar. En les espècies gregàries, si el meconi és de color vermell, quan neixen molts adults de cop i al mateix indret, apareixen nombroses taques rojes a terra, les quals constitueixen les anomenades pluges de sang; això passa, per exemple, en els nimfàlids Inachis io i Aglais urticae. Si la crisàlide té embolcall pupal sense forat de sortida, el meconi conté substàncies càustiques per a dissoldre’l. Si no hi ha capoll, el naixement es pot endevinar perquè l’exúvia deixa veure les ales, plegades però ja formades i acolorides.
A la metamorfosi, s’ha comprovat que, en tots els processos de transformació, hi ha un elevat grau de compartimentació morfològica i funcional. Si hom extirpa a una eruga uns discs imaginals determinats, la crisalidació es produeix igual, i neix una papallona a la qual manca un òrgan, però capaç de viure i reproduir-se perfectament.
Una vegada fora de la crisàlide, l’imago estén les ales injectant hemolimfa per les nervacions alars i aire pels conductes traqueals. Després es col·loca en un lloc assolellat per tal d’assecar-se les ales i endurir-les, i així poder volar; el constant ús d’aquesta activitat és molt característic de l’ordre.
Comportament i defensa
Per la mateixa naturalesa interactiva i globalitzadora del comportament, moltes pautes de conducta ja han estat descrites en els apartats anteriors, i d’altres ho seran en els posteriors; aquí només tractarem les que no s’escauen en parlar dels altres temes.
Els mitjans de defensa individual
Els lepidòpters asseguren la seva supervivència individual mitjançant unes pautes de comportament de caire general i d’altres més concretes, que es desencadenen en el moment de perill. Les primeres són comunes a grans grups, mentre que les segones les solen tenir un nombre més restringit d’espècies; ambdues, però, es basen en una determinada estructura morfològica. La defensa és, doncs, el resultat d’una compaginació de la morfologia i la conducta, que han evolucionat conjuntament.
La majoria de lepidòpters, tant en estat adult com d’eruga, són d’activitat nocturna, cosa que dificulta la localització als depredadors. Un altre hàbit general és el d’amagar-se, les larves, dins de l’aliment, i, els adults, a la fullaraca del terra. Algunes erugues, per a augmentar la protecció, torcen les fulles, com les dels tortrícids (d’aquí el seu nom); d’altres excaven galeries dins la planta (endòfites) o a terra (endogees). Els microlepidòpters, si els amoïnen, es deixen caure, encongits, i es queden quiets per tal de semblar encara més petits i, així, passar desapercebuts o resultar menyspreables per a molts depredadors. Alguns practiquen molt bé la immobilització, i es fan el mort d’una manera tan convincent, que no resulten abellidors. En canvi, els lepidòpters grossos tendeixen a obrir les ales per tal de dissuadir els petits depredadors. Les espècies de vol ràpid emprenen la fuga sempre que ho consideren factible.
Les femelles realitzen habitualment la posta als llocs més arrecerats; les que tenen l’ovopositor allargat ponen els ous dins l’escorça dels arbres, els fruits, les fulles, etc. i d’altres cobreixen la posta amb pèls o substàncies protectores. La major part d’erugues tenen pèls que, com a mínim, dificulten l’operació d’engolir-les als petits depredadors. Algunes, com les de molts nimfàlids, tenen pèls transformats en espines, i d’altres són urticants (com les processionàries). Assenyalarem que l’embolcall pupal pot significar una eficacíssima protecció per tal com alguns són molt durs, d’altres elaborats amb estelles de fusta, i d’altres són urticants. Recordem que moltes papallones, a més, crisaliden sota terra.
Pel que fa a les pautes de comportament més específiques, podem assenyalar l’aspecte espaoridor d’algunes erugues. Per tal d’espantar més l’intrús solen adoptar postures determinades; en el cas dels esfíngids, aixequen el cap i el tòrax (i d’això els ve el nom, car semblen una esfinx). Algunes erugues tenen glàndules repugnatòries: els licènids i limàntrids les tenen en posició dorsal; els nimfàlids, noctúids i notodòntids, en posició ventral, i llancen el líquid a distància. La papallona rei (Papilio machaon) i d’altres espècies de la seva família, tenen una glàndula postencefàlica evaginable, la qual és impregnada de pudent àcid butíric, que projecten contra l’intrús. Hi ha erugues, com el cuc de la col (Pieris brassicae), que regurgiten un suc verdós, agre i càustic, que pot resultar mortal per a alguns depredadors.
Quant a l’estat adult, indicarem tot seguit els mitjans específics de defensa més coneguts. L’autohemorràgia és un sistema usat pels àrctids, com Arctia caja, i consisteix a secretar hemolimfa tòxica i repel·lent a través del tegument mitjançant uns solcs que obren a voluntat. Els mateixos àrctids, els noctúids, i moltes altres famílies de nocturnes que, en posició de repòs són ben camuflats, sorprenen i poden espantar els depredadors aixecant les ales anteriors, i deixant al descobert les posteriors, que són de colors vius. Quelcom semblant passa amb altres espècies, algunes diürnes, en mostrar els ocel·les, de gran similitud amb els ulls dels vertebrats. Hi ha casos més complets, com el gènere exòtic Caligo, que en posició posterior invertida sembla tota ella la cara d’un mussol (per això hom les coneix com a papallones mussol).
En el camp de l’acústica, assenyalarem la papallona de la mort (l’esfíngid Acherontia atropos) que, en ser inquietada, emet una fressa i uns xiulets molt clars injectant aire per l’espiritrompa i fregant entre ells els darrers segments abdominals. Un sistema més complex és el dels noctúids, els quals, quan capten (des de més de 30 m) els ultrasons d’ecolocalització dels rats-penats que els persegueixen emeten també ultrasons per tal de confondre’ls.
Cripsi
Dins l’homotípia, el camuflament més comú en els lepidòpters és l’homocromia, que tenen la majoria d’espècies, les quals imiten escorça, fulles, etc. L’homomòrfia ja és més rara, però hi ha casos de semblança molt accentuada. Així, tenim el lasiocàmpid Gastropacha quercifolia que simula molt bé un piló de fulles seques, i el drepànid Cilix glaucata, que sembla un excrement d’ocell. Les espècies del gènere exòtic Kallima són un duplicat perfecte de les fulles dels arbres, però, a més, tenen un fenomen de coevolució entre elles.
Per a camuflar-se millor, les ales anteriors solen cobrir les posteriors. És així en moltes espècies nocturnes, algunes de les quals, a més, pleguen les ales en forma de teulada (noctúids) per a eliminar les ombres que augmentarien el contrast. Els geomètrids, però, disposen les ales aplanades sobre el substrat i molt poc sobresortints. Molts microlepidòpters envolten amb les seves ales la fina tija sobre la qual se situen verticalment per a resultar menys patents. Tanmateix, els ropalòcers pleguen les ales verticalment; aleshores, la part vistent és l’anvers, mentre que el revers és críptic, sobretot el de les ales posteriors, que pot cobrir les anteriors. Alguns ropalòcers, en detectar perill, segueixen el moviment de l’intrús mantenint-se en posició frontal per tal que la superfície exposada sigui la mínima possible.
Tots els casos indicats fins ara són d’imagos. En les erugues també és molt habitual l’homocromia, amb ornamentacions molt críptiques amb la planta hoste. Situacions d’homotípia les trobem en els geomètrids; els coleofòrids i psíquids utilitzen l’enginyós sistema, emprat també per altres grups, de no modificar la morfologia, sinó de cobrir-se directament de fulles i branques amb la qual cosa ja no hi ha semblança, sinó identitat.
Protecció a nivell d’espècie
Un dels casos més coneguts de protecció a nivell d’espècie és el de les erugues processionàries del pi (Thaumetopoea pityocampa). Aquestes, a més de ser molt tòxiques fins i tot sense ésser ingerides, a causa dels pèls urticants, són gregàries. Aquest gregarisme és dels més accentuats, car neixen de postes agrupades, i fabriquen grossos nius de seda, dins dels quals viuen les erugues. Quan surten, van seguint un fil de seda (fent una processó) i només se separen per a alimentar-se. Després de menjar, tornen totes juntes al niu reseguint els fils de seda. Fins i tot, crisaliden conjuntament, i les crisàlides són urticants i subterrànies. Cada eruga té 700 000 espines finíssimes untades amb una secreció urticant. Aquesta secreció es converteix en un polsim que provoca al·lèrgies i irritacions epidèrmiques (fins i tot sense necessitat de tocar l’eruga) que poden ser greus, per exemple, a les mucoses oculars. Si s’ingereixen provoquen greus intoxicacions que poden ser mortals (fins i tot per a l’home). Aquest sistema de defensa, combinació del tipus individual directe i de la protecció a nivell d’espècie, accentuada pel gregarisme i pels nius infestats de pèls urticants, és dels més eficients que es coneix. Potser per això no necessita colors aposemàtics.
En estat adult, els lepidòpters habitualment no són tòxics. En conseqüència, les coloracions i dissenys aposemàtics no són gaire estesos. Tanmateix, es donen en els àrctids, els zigènids, i en els exòtics helicònids i nimfàlids de la subfamilia dels danaïns (Danainae). En els adults d’aquestes famílies es dona el mimetisme mullerià. El cas més evident, i present a casa nostra, el tenim en els cercles sinaposemàtics del gènere Zygaena, totes les espècies del qual semblen portar el mateix uniforme. Aquest mimetisme comporta una protecció de l’espècie molt eficient i és el resultat d’una coevolució. Als Països Catalans també trobem casos més complicats, com el de Z. ephialtes, espècie polimòrfica que pertany a dos cercles sinaposemàtics diferents.
En correspondència als casos de mimetisme mullerià que hem citat, trobem els casos de mimetisme batesià. Per exemple, els helicònids i danaïns tòxics són mimetitzats per papiliònids i pièrids inofensius, i de vegades assoleixen una similitud sorprenent que arriba a fer difícil la distinció. Però aquests casos es donen en altres indrets. Als Països Catalans també trobem mimetismes molt clars i ben aconseguits: els dels sèsids i alguns esfíngids, que semblen abelles i altres himenòpters.
El vol
És ben sabut que el mitjà de trasllat més important dels lepidòpters és el vol; l’utilitzen per a alimentar-se, per a buscar la parella, per a fugir dels depredadors, per a dispersar-se, per a migrar, etc. Pel fet d’ésser poiquiloterms, els lepidòpters necessiten arribar a una temperatura mínima abans d’emprendre el vol. Les espècies diürnes capten la calor amb les ales en prendre el sol; els esfíngids, i altres famílies d’activitat nocturna, fan vibrar les ales per tal de generar calor.
Tots els lepidòpters tenen un vol de tipus sincrònic, amb una freqüència de batec sempre inferior a 100 Hz; però, dins d’aquest tipus, hom pot distingir tres maneres de volar: el vol planat, que és vol a vela, planejant amb les ales esteses i quietes, que per a ésser efectiu, necessita una proporció superfície alar / pes elevada, com en els ropalòcers; el vol batut, que és un moviment actiu de les ales, però de baixa freqüència (fins a uns 20 Hz), i és el més general en els lepidòpters i l’obligat en les espècies de cos gros (moltes famílies de nocturnes que pesen massa per a poder planejar); i el vol vibrat, que consisteix en un moviment ràpid de les ales, a més de 20 Hz, cosa que permet més acceleració i maniobrabilitat, i poden arribar a parar-se en ple vol; per això les ales han de ser estretes i fermes. Aquest darrer tipus de vol l’usen pocs lepidòpters; entre els que aconsegueixen una freqüència més alta hi ha el bufaforats (Macroglossum stellatarum) i els sèsids: s’ha calculat que Sesia apiformis arriba a efectuar 96 batecs per segon.
Moltes espècies usen alternadament el primer i el segon tipus; en canvi, els que usen el tercer ho fan exclusivament. Independentment de la freqüència de batec d’ales, els lepidòpters poden assolir velocitats de vol més o menys grans, sense relació directa amb la freqüència. Una velocitat mitjana seria la de la papallona de la col: 13 km/h. Les espècies més ràpides les trobem entre les nocturnes i les crepusculars grans, com els esfíngids, que tenen un cos molt aerodinàmic i assoleixen una velocitat de 60 km/h (fixem-nos en què significa això per a un animal tan petit).
Les migracions
Totes les espècies, en els estats d’eruga i d’imago, en realitzar les activitats habituals de la seva existència, es dispersen més o menys; això difícilment comporta un increment de l’àrea de distribució. Moltes espècies de lepidòpters realitzen migracions locals (hom anomena migració al trasllat dels adults d’un mínim d’uns quants quilòmetres), és a dir, restringides a zones on l’espècie ja viu i completa el cicle biològic; la finalitat d’aquestes migracions seria disminuir la consanguinitat i mantenir elevada la capacitat adaptativa de l’espècie. Però existeix un altre grau de migració: els llargs desplaçaments de centenars i, de vegades, milers de quilòmetres. Aquest fenomen permet colonitzar noves àrees, on es poden formar subespècies i, fins i tot, espècies noves, sempre que es produeixi un aïllament posterior d’alguna àrea perifèrica. Les migracions les poden efectuar exemplars aïllats (com és el cas de molts esfíngids), però el més normal és que les efectuïn força exemplars a l’hora, encara que rarament en grups molt compactes. Hom ha constatat l’existència de grups de milions d’individus del noctúid Autographa gamma en migració.
A les nostres latituds, és habitual que, en una espècie migradora, els adults nascuts a la primavera volin cap al N, realitzin allà un cicle complet, i els nous imagos, a l’estiu o la tardor, retornin vers el S (més càlid), a passar l’hivern. Això és el que fa la papallona monarca (el nimfàlid Danaus plexippus), amb la particularitat que el viatge d’anada i tornada el fan els mateixos individus, cosa molt rara; però no solament per això és una de les espècies migradores més espectaculars, sinó també per la seva grandària i pels llargs viatges que efectua (va del Canadà a Mèxic). És impressionant veure, als seus llocs d’hivernada de Mèxic, els milions d’exemplars que cobreixen el terra i els arbres, i arriben a donar color al paisatge. A més, alguns individus travessen l’Atlàntic, i arriben fins a les illes Canàries i a la península Ibèrica. Hom l’ha vista reposar sobre la mar en calma, amb les ales esteses, abans de reprendre el vol. Les migracions són afectades per la direcció dels vents, ja que els lepidòpters són organismes fàcilment transportables pel corrent. La rotació de la Terra fa que les grans migracions siguin cap a l’E. Per això la papallona monarca no travessa l’Atlàntic cap a Amèrica.
Normalment, els individus que migren són ben alimentats, però de vegades la migració ve determinada per la manca d’aliment al lloc de naixement, bé per escassetat de la planta nutricia, bé per superpoblació del lepidòpter. Un altre factor que pot afavorir les migracions és una variació climàtica, ja que això permet la colonització de llocs on normalment el clima ho impedeix. Uns dels llocs idonis per a l’observació de migracions són: el litoral (on es troben espècies procedents del mar, perquè s’aturen a descansar) i els passos de muntanyes. Els depredadors ho saben i s’hi concentren, i per això, els danaïns i els helicònids, que migren en bandades i serien una presa massa fàcil i atraient, tenen sistemes de protecció a nivell d’espècie (toxicitat o mal gust).
Fototropisme
La divisió dels lepidòpters en nocturns i diürns, bé que molt usada, és força confusa, sobretot pels nombrosos casos d’espècies d’activitat diürna que pertanyen a famílies de nocturnes (per exemple, el bufaforats). De vegades, la femella és nocturna i el mascle diürn, com el paó de nit (Pavonia pavonia) i el lasiocàmpid Lasiocampa quercus. El que sí que es pot assegurar és que tots els ropalòcers del nostre país són heliòfils (d’activitat diürna), i també els zigènids; i gairebé tots són termòfils. Alguns ropalòcers exòtics (satirins i hespèrids) són crepusculars i fins i tot nocturns.
Una de les facetes del comportament més sorprenents dels lepidòpters és el fototropisme positiu que tenen gairebé totes les espècies d’activitat nocturna o crepuscular. La seva atracció per la llum és més accentuada en la franja dels ultraviolats. Sovint s’apropen al focus de llum donant voltes, i tot fa pensar que ho fan així per a conservar l’angle d’apropament determinat per una franja lateral de facetes dels ulls compostos; de vegades, van donant voltes a la llum fins a cremar-se les ales, d’altres s’aturen al costat i algunes no marxen fins hores després de l’alba. Potser no cal cercar una explicació d’aquesta conducta fototròpica, de la qual s’han fet nombroses interpretacions, car una llum en plena nit és un fenomen totalment estrany en la vida dels lepidòpters, i en conseqüència, l’evolució no té perquè haver previst una resposta especialment adequada. En general, la instal·lació de llums permanents té conseqüències negatives per a la comunitat de lepidòpters, per tal com són un factor distorsionador a la vida de l’ecosistema, i fa que la pressió de depredació augmenti molt sobre els grups de papallones que es concentren a l’entorn dels focus.
En les erugues també es donen fototropismes. Per exemple, les del grup Vanessa (nimfàlids) es traslladen en direcció solar i, si es mouen, recuperen l’orientació. D’altres espècies passen d’un fototropisme positiu a un de negatiu segons que hagin menjat o no.
Ecologia
A l’inici d’aquest mateix capítol ja hem qualificat els lepidòpters des del punt de vista ecològic. Parlarem, ara, del paper que juguen en el segon nivell tròfic dels ecosistemes terrestres, de com obtenen l’energia del primer nivell (alimentació), i de com la cedeixen als macròfags carnívors (depredació). Són, per tant, organismes de ràpida multiplicació i de vida curta, que segueixen clarament l’estratègia ecològica de la r i són, a més, típiques preses dels predadors. Hi ha casos, també, que s’aparten d’aquest model general i se situen a d’altres nivells o a línies colaterals de transferència d’energia. Així mateix, tractarem de la influència dels factors ambientals en la biologia dels lepidòpters i dels desequilibris que ocasionen. Un fet que hom ha posat de relleu és la gran dependència de les poblacions de lepidòpters en relació a la vegetació; és tan estreta que hom ha detectat una elevada correlació entre la diversitat de les poblacions de lepidòpters i la de la coberta vegetal. Parlarem separadament de l’alimentació de les erugues i dels adults perquè, malgrat constituir una mateixa espècie, ecològicament tenen relacions totalment diferents.
L’alimentació de les erugues
Després d’un estudi meticulós de l’alimentació de les erugues hom podria arribar a la conclusió que mengen de tot. Efectivament, el ventall alimentari és ample, cosa que és previsible si hom considera l’elevat nombre d’espècies de lepidòpters existent, i el seu èxit ecològic. La potència de les seves mandíbules és palesa en el noctúid Brithys crini, que perfora el plom. Tanmateix, hom pot dir que les preferències es decanten molt clarament per les plantes, i la majoria són polífagues. Dins de les fitòfagues, la major part són fil·lòfagues (mengen fulles), però d’altres són antòfagues (mengen flors) i d’altres s’alimenten dels brots; molts microlepidòpters mengen llavors (carpòfags). Força famílies tenen erugues endòfites, i destaquen les minadores, típiques dels microlepidòpters més petits, que ingereixen el parènquima de les fulles i produeixen galeries; d’altres s’alimenten de suro, arrels i tota mena de fruits; altres viuen a l’interior dels cactus i fins i tot algunes són cecidògenes i produeixen les hipertròfies de teixits vegetals anomenades cecidis o gal·les. Veiem, doncs, que no hi ha cap part de la planta que se salvi de l’atac dels lepidòpters. Pel que fa als grups vegetals, el de les plantes superiors (fanerògames) és el preferent, sense despreciar les paràsites ni les aquàtiques; però hi ha erugues micòfagues i d’altres que mengen molses, líquens (com els nolins i els àrctids), cues de cavall (Equisetum) i, fins i tot, un grup poc explotat pels insectes: el de les falgueres.
Deixant a part les espècies fitòfagues, passarem a les que se situen a altres graons de la cadena tròfica. Aquest és el cas de les sapròfites que aprofiten les matèries en descomposició, tant de teixits vegetals com animals putrefactes, detritus (detritòfagues) i directament excrements (copròfagues). Entre les erugues que mengen teixits animals destaquen els tinèids, que ataquen fonamentalment pèls, plomes i materials manufacturats com llana, flassades, estores, tapisseria i vestits i robes en general, inclòs el cuir, la pell i substàncies còrnies. El poll o tinya de la cera (el piràlid Galleria mellonella) entra als ruscs de les abelles i s’alimenta de la cera. D’altres de la mateixa família i alguns ecofòrids són comensals de l’home en ingerir farina, mantega, greix, etc.; hi ha espècies que es mengen els insectes de les col·leccions i altres animals dissecats, així com les plantes seques dels herbaris, el paper i el cartró. Experimentalment, hom ha confeccionat molts tipus de pastes nutritives, per a alimentar-les en captivitat.
Tots els casos citats tenen en comú l’estratègia ecològica tròfica: la microfàgia, en el sentit que esmicolen l’aliment en un procés que no demana gaire atenció. Seguidament, indicarem algunes espècies macròfagues, de classificació dubtosa o situades en línies de transferència d’energia separades de la principal. D’entre les espècies carnívores destaquen uns licènids tropicals, que són autèntics depredadors d’altres insectes i petits animals; algunes fins i tot es devoren entre elles. Pel que fa al parasitisme, també trobem exemples entre els lepidòpters: hi ha gelèquids que xuclen el suc que desprenen les cotxinilles, i el piràlid Sthenauge parasitus, que pon els ous sobre erugues d’altres lepidòpters (satúrnids).
Finalment, indicarem els casos de comensalisme i simbiosi. Hi ha erugues que viuen als nius de tèrmits, formigues, aranyes, abelles, vespes i, fins i tot, ocells i mamífers. Atesa la llargada de la llista, tan sols exposarem un cas, potser el més important: la relació amb les formigues. Hi ha licènids mirmecòfils que, en alguna fase larval, poden deixar de menjar la planta hoste i entrar als formiguers, on s’alimenten dels materials de reserva d’aquests. El cas extrem el trobem en algunes erugues que tenen glàndules que secreten un suc que agrada a les formigues, les quals les transporten per les galeries tot alimentant-les fins que crisaliden. Però als Països Catalans viu un licènid, Maculinea arion, que és mirmecòfag.
L’alimentació dels adults
Si les erugues destrueixen les plantes, els lepidòpters adults col·laboren al seu desenvolupament, freqüentment de manera decisiva, per mitjà de la pol·linització. Tots els imagos viuen de les reserves acumulades per l’eruga, però molts prenen un suplement alimentari, que sol ser nèctar de les flors. Com que es tracta de sucres, sense proteïnes ni greixos, és una dieta incompleta, i representa, més que una alimentació, l’adquisició d’un combustible per tal de reposar energies per a poder seguir volant. De fet, adults que habitualment s’alimenten, mantinguts en captivitat sense prendre res, viuen i es reprodueixen perfectament, si bé es moren abans que els altres. Aquesta situació es pot interpretar com una reminiscència d’ancestrals necessitats, o com a l’afavoriment de la reproducció de la planta nutricia, assegurant l’aliment de l’eruga (en els casos en què sigui la mateixa).
En xuclar el nèctar, s’adhereixen a l’adult grans de pol·len, a l’hora que se’n desprenen dels que ja porta d’altres flors. Així es produeix la pol·linització. Aquesta resulta tan important per a algunes plantes, que s’ha produït una coevolució entre elles i els lepidòpters. Per la seva banda, els imagos s’asseguren una font d’energia de difícil accés per a la majoria d’insectes, ja que els nectaris poden situar-se a força centímetres de fondària; l’adult corresponent ha anat desenvolupant l’espiritrompa adequada. Per tal d’optimitzar aquesta col·laboració, la flor s’ha anat fent més conspicua, amb formes, colors i olors més diferenciades. Així, en distingir bé la flor, l’adult visita amb més probabilitat flors de la mateixa espècie, i deixa el pol·len als estigmes adequats; sol guiar-se per esquemes i dissenys, de vegades de llum ultraviolada, que nosaltres no podem veure, però que la flor posseeix. Hom comprova que reconeixen i intenten libar dibuixos esquemàtics pintats. Sovint, una espècie visita diversos vegetals, que normalment són de la mateixa família.
Els adults d’algunes espècies (molts nimfàlids) són atrets per l’olor de les fruites madures, la saba i altres productes vegetals. Els micropterígids mengen amb les mandíbules (cas excepcional) els grans de pol·len. Però també hi ha espècies que aprofiten productes d’origen animal, com ara líquids en putrefacció, excrements i altres substàncies que desprenen els animals: el líquid ensucrat dels pugons i les secrecions greixoses del mandrós (mamífer edentat) són exemples de comensalisme. La papallona de la mort o esfinx de la calavera (Acherontia atropos) xucla grans quantitats de mel (més que el seu propi pes); dins el rusc emet un so que imita el de la reina per tal d’eludir les abelles, però les picades d’aquestes poden arribar a matar el lepidòpter.
Depredadors
Tractarem ara dels animals als quals els lepidòpters transfereixen l’energia. Entre els depredadors típics dels lepidòpters, és a dir, els macròfags carnívors, destaquen alguns vertebrats, i altres insectes. Entre els petits vertebrats sobresurten els ocells, grans caçadors de papallones en totes les seves fases; s’ha comprovat que, en alguns casos, ells sols són els responsables que més del 95 % dels ous de papallona no arribin a adults. Els mamífers insectívors també són importants: els rosegadors (rates, talps, etc.) detecten moltes erugues i crisàlides amb l’olfacte; els rats-penats localitzen els adults de les espècies nocturnes i crepusculars en ple vol amb el radar. Altres vertebrats més grossos, rèptils (llangardaixos, camaleons i altres) i amfibis (granotes, gripaus, etc.), també ataquen els lepidòpters. Tampoc s’ha d’oblidar l’acció dels grans herbívors que ingereixen juntament amb l’herba quantitats importants de lepidòpters en estat larval. Encara que no d’una manera tan espectacular com els vertebrats, els insectes, pel seu gran nombre, incideixen enormement sobre les poblacions de lepidòpters. Entre els coleòpters remarcarem el conegut cas del caràbid Calosoma sycophanta, gran caçador d’erugues; quan hi ha hagut plagues de Lymantria dispar a Catalunya, les poblacions d’aquest caràbid han augmentat molt, i han constituït un factor estabilitzador. És habitual, en aquests casos, veure’ls empaitar les erugues amb gran agressivitat i, sovint, matar-les sense ingerir-les.
Altres insectes depredadors d’erugues són els planipennis (gènere Chrysopa) tant en estat larval com adult, heteròpters, dípters (sírfids), mecòpters (gènere Panorpa o mosca escorpí), ortòpters, ensífers, dictiòpters, etc.; entre aquests últims destaca el pregadéus (Mantis religiosa), que ataca també els adults a l’aguait. Els himenòpters són molt importants: algunes formigues injecten verí a les erugues i després els xuclen l’hemolimfa. Però altres formigues, així com les vespes, mengen directament les erugues tot utilitzant les mandíbules. Entre els artròpodes no insectes, els àcars buiden els ous de la substància nutritiva; les aranyes cacen els adults amb la tela, etc.
Paràsits i malalties
Els lepidòpters, sobretot en estat larval, i especialment els que tenen mitjans de defensa efectius contra el depredadors típics, són molt atacats per tot tipus de paràsits: grossos (alguns himenòpters) i petits (la majoria d’insectes) fins i tot microscòpics (microbis), endoparàsits (la majoria) i ectoparàsits (àcars), específics o no, solitaris (alguns himenòpters grossos) o gregaris (la majoria), i fins i tot hi ha alguns casos de poliembrionia (calcidòids, bracònids i altres himenòpters), hiperparasitisme, etc. D’entre els poliembrionaris, hom pot destacar un cas extrem de gregarisme: dins una eruga de Chrysodeixis chalates (noctúid) hom trobà 2000 larves del gènere Litomastix (calcidòid encírtid).
D’entre els himenòpters destaquen, de més grossos a més petits: icneumònids, bracònids i calcídids (de menys d’1 mm). Hi ha bracònids tropicals que, amb l’oviscapte de més de 10 cm, perforen l’escorça dels arbres i ponen els ous sobre les erugues endòfites. Aphiantax té un oviscapte que fa 14 vegades la longitud del cos. Entre els dípters hi ha nombrosos paràsits, però tenen sistemes semblants als dels himenòpters; en destaquen els taquínids (paràsits forçosos). En alguns casos, fan la posta sobre la planta nutricia de l’eruga i els ous s’adhereixen a la seva cutícula. Els paràsits microscòpics se solen estudiar a part, i la seva presència provoca malalties; les més habituals són les bacteriosis i les virosis, que solen provocar pèrdua de gana i de mobilitat a l’eruga, la qual acaba per morir i liqüar-se. El procediment més normal d’infecció és per ingestió de l’aliment. El bacteri més conegut és Bacillus thuringiensis, que entra en forma d’espores i passa per contagi d’una eruga a l’altra. Els virus entren en forma de grosses partícules virals infeccioses, que per a entrar també poden aprofitar lesions cuticulars. Les micosis es distingeixen fàcilment quan fructifica el miceli. Entre els fongs més comuns es troba Penicillium, Aspergillus, etc., i altres que són mortals com Beauveris, Metarhizium, etc. En canvi, les malalties ocasionades pels protozous solen ésser menys patògenes (pèrdua de fecunditat i longevitat).
Influència dels factors ambientals
Totes les fases del cicle biològic dels lepidòpters són afectades pels múltiples factors ambientals. Ens referirem als més importants.
Pel que fa a la temperatura, és el d’efecte més directe, ja que arriba a alterar la morfologia (color i dissenys alars). Cada espècie té el seu interval òptim de temperatura, en el qual l’activitat i la supervivència és màxima; l’òptim es refereix a la temperatura mitjana, ja que les variacions han d’existir per induir els canvis i mantenir la taxa de fertilitat. Amb temperatures superiors, la velocitat del cicle augmenta, els exemplars surten més petits i foscos i fins es pot incrementar el nombre de mudes, però, a partir d’un nivell (de 40 a 50° C), apareix el risc de mort per coma calòric. Si no es perllonga gaires hores, alguns ous aguanten més de 60° C. La resistència dels lepidòpters a altes temperatures o a gelades varia segons l’espècie, la fase en què es trobi i la rapidesa del canvi tèrmic, que determina que es pugui adaptar a la variació o no. Per això, quan es troba en diapausa, té el màxim de resistència. Malgrat que prefereixen normalment ambients càlids, és sorprenent la resistència d’algunes espècies adaptades al fred. En aquests casos, és essencial el perfecte funcionament de la regulació hídrica dels teixits perquè puguin actuar les substàncies anticongelants. Hi ha espècies que es deshidraten i així poden romandre congelades durant força temps; quan es descongelen, es rehidraten i recuperen l’activitat normal. Hom ha observat adults volar a prop dels 20° C sota zero. En estat larval, s’han constatat els –50° C, i alguns ous suporten els –80° C.
La humitat, juntament amb la temperatura, és el paràmetre que influeix més en la vida dels lepidòpters, sobretot perquè fa variar les característiques de la cutícula. Per exemple, un problema típic és que, si no hi ha suficient humitat, durant la muda l’exúvia no es desprèn del tot, cosa que pot matar l’eruga. Una sequera pot ocasionar una gran mortalitat perquè la deshidratació forçada dels òrgans actius és fatal. L’embolcall pupal és decisiu per a mantenir el mínim d’humitat. Bé que alguns ropalòcers crisaliden a ple sol, les erugues, sobretot les que no fan capoll, solen cercar un lloc ombrívol i humit per a crisalidar. L’òptim higromètric se situa entre el 60 i el 70 %. Pel que hem vist, ja es pot deduir que els lepidòpters són més sensibles al descens d’humitat que a les pujades. Fins i tot s’ha observat un canvi en el comportament si baixa molt la humitat: algunes erugues arriben a menjar-se entre elles per tal de mantenir el mínim hídric intern. En estat larval, però, un excés d’humitat tampoc no és bo, car afavoreix la instal·lació de fongs.
L’altitud, el fotoperíode, el tipus de terreny, la concentració d’oxigen, la pluviometria, el vent i el clima en general influeixen en la vida dels lepidòpters; però és impossible detallar l’efecte de tots especialment per la diversitat de les respostes i perquè uns paràmetres vénen condicionats per d’altres. A més, els factors interaccionen, i produeixen un efecte diferent segons la magnitud dels altres. Per exemple, l’òptim de temperatura varia si es modifica la humitat, i a la inversa. Una eruga que ha perdut l’activitat per una temperatura baixa, pot recuperar-la si augmenta la humitat. També s’observen canvis en el comportament, com ara en els fototropismes, etc.
El lepidòpter s’informa de l’època de l’any captant diferents factors ambientals, com la temperatura i el fotoperíode. Els canvis solen provocar el pas d’una fase a l’altre: un descens de la pressió atmosfèrica desencadena el naixement dels imagos.
Plagues
Les plagues són una conseqüència del desequilibri ocasionat als ecosistemes. En condicions normals, les poblacions de lepidòpters, com les de tots els éssers vius, són controlades pels depredadors i els paràsits, pels factors ambientals, etc. Una alteració d’aquests controls, que en altres animals pot comportar lleugeres oscil·lacions en el nombre d’individus, en els lepidòpters pot provocar autèntiques explosions demogràfiques. I això és degut bàsicament a dues característiques fonamentals: la ràpida successió de les generacions i l’estratègia de quantitat que usen, que es tradueix en una elevada taxa de reproducció. Una posta normal té alguns centenars d’ous, però hi ha espècies que en ponen més de 4000. Això fa que la pressió dels depredadors hagi d’ésser constant. La disminució, ni que sigui transitòria, d’aquesta pressió, comporta ràpidament un increment de la població. La majoria de lepidòpters tenen molts depredadors, que els provoquen una elevada taxa de mortalitat, que compensa la de natalitat. Però si disposa d’uns mitjans de defensa molt efectius, el ventall de depredadors és poc diversificat, car han d’estar específicament adaptats per superar les defenses. Aquest és el cas de la processionària del pi, que té pocs enemics: diversos ocells, com el cucut reial (Clamator glandarius) i formigues del grup Formica (com F. rufa), a part d’alguns paràsits microscòpics. Aleshores, quan fallen aquests controls, no n’hi ha d’altres substitutoris. A més, algunes espècies varien el nombre d’ous de cada posta segons la quantitat d’aliment que ha trobat l’eruga. S’ha comprovat que Lymantria monacha pot duplicar-lo.
A part això, hi ha factors artificials que afavoreixen les plagues de lepidòpters: els monocultius, les millores genètiques (solen augmentar la producció però també disminuir les defenses de la planta) i qualsevol homogeneïtzació dels conreus, i també les alteracions de l’equilibri dels ecosistemes. Aquestes les poden provocar els herbicides, les replantacions d’arbres no autòctons, els mateixos insecticides, i l’immens volum d’intercanvi de productes vegetals a escala mundial, cosa que ocasiona l’extensió de greus plagues a llocs on no tenen enemics naturals pel fet de no haverhi viscut mai. Podem posar-ne un exemple: ja fa més d’un segle, a Massachusetts, a un científic se li escaparen uns exemplars de Lymantria dispar que procedien d’Europa; l’espècie introduïda es convertí en el principal defoliador dels boscos nordamericans i la plaga, que no s’ha pogut eliminar, actualment continua provocant gravíssims flagells. Lamentablement, podríem explicar molts altres casos com aquest.
Una característica de les plagues dels lepidòpters és que solen ser defoliadores. Sovint provoquen defoliacions que no matarien l’arbre, però en debilitar-lo per falta de fotosíntesi, acudeixen altres insectes, com els escolítids (coleòpters), atrets per les substàncies que l’arbre desprèn; en aquest sentit, els escolítids són temibles perquè les plagues que ocasionen són de les més terribles que es coneixen: la d’Hondures, que durà tres anys, va fer desaparèixer dos milions i mig d’hectàries de bosc (arribaren a morir 162 000 arbres diàriament). Bé que potencialment tots els lepidòpters poden ocasionar plagues, la majoria no ho fan mai. La lluita contra les plagues de lepidòpters comença, a escala preventiva, fent el contrari del que hem dit que les ocasiona o afavoreix. Malgrat això, les espècies polífagues són pràcticament impossibles d’eliminar, ja que si se’ls impedeix d’actuar sobre una planta, passen a menjar-ne una altra. L’acció directa d’insecticides (DDT i altres organoclorats) significà, els anys quaranta, una revolució en els mètodes, i resultà molt més efectiu que els sistemes tradicionals. Però, com ja hem apuntat, el mateix insecticida pot ocasionar desequilibris que desencadenin noves plagues. Per tal d’evitar això actualment es fumiga amb espores de Bacillus thuringiensis, que són tòxiques només per a les erugues de lepidòpter. Normalment l’eficàcia de la lluita biològica és baixa, i sempre perillosa pel mateix fet d’introduir noves espècies.
Actualment es comencen a usar les feromones sintètiques per a atrapar els mascles, impedint la reproducció. Indiscutiblement, no es pot trobar un sistema més específic i sense efectes secundaris, i és el millor procediment amb els lepidòpters que tenen aquest tipus d’atracció. Malgrat tot, els lepidòpters són un grup tan ben adaptat a les condicions actuals de la terra, i són tan versàtils, que sempre troben refugis i maneres d’eludir l’acció humana. Per això, avui en dia, més que mantenir la idea d’acabar amb les plagues, es pretén la lluita integrada, és a dir, estudiar profundament la biologia del lepidòpter i buscar tots els sistemes per combatre’l, aplicar els que siguin més efectius, i, així, fer recular la plaga a un grau en què el resultat sigui econòmicament rendible.
Distribució geogràfica
En particular, la distribució de cada espècie de lepidòpters depèn de molts factors. En principi, depèn del lloc on s’ha originat l’espècie, així com de les múltiples vicissituds que hagi sofert al llarg de la seva història evolutiva: canvis geològics, climàtics, pressió d’espècies noves que entren en competència, dels depredadors, etc. Des del punt de vista paleozoogeogràfic, els troncs evolutius de la major part dels lepidòpters dels Països Catalans tenen el seu origen a la regió asiàtica oriental. Han arribat seguint rutes europees (pel N de la mar Tetis) o africanes (pel S).
També tenim algunes espècies derivades de troncs originats al continent africà. Concretament, el 70 % de la fauna lepidopterològica catalana ha arribat seguint la ruta eurasiàtica o la mediterrània; la resta ha seguit les rutes atlàntica i etiòpica.
A partir del lloc on s’ha originat cada espècie, els lepidòpters poden dispersar-se mitjançant procediments voluntaris (migracions) o, més habitualment, involuntaris: portats pels corrents de l’aire, pels animals (entre el pèl, per exemple) i, recentment, té gran importància el comerç mundial de plantes útils per a l’home. Molts d’aquests trasllats són factibles en fase d’ou o crisàlide.
Les espècies monòfagues veuen limitada la seva àrea de distribució per la de la planta nutricia. Sovint, la del lepidòpter és més reduïda perquè els vegetals s’adapten a llocs inhabitables pels lepidòpters. Però el més normal és que les barreres a la dispersió les constitueixin les serralades, els mars, els grans llacs i les zones gelades, els deserts i els canvis de clima. Els rius solen servir de tascons d’entrada de lepidòpters a noves zones, i actuen de pont per a superar les barreres. La presència d’una altra espècie que competeix més eficaçment pel mateix substrat alimentari també és una barrera, bé que no geogràfica, i és l’origen de les típiques poblacions al·lopàtriques que se substitueixen mútuament a una zona i l’altra. L’acció humana (amb la caça o, indirectament, amb la degradació ambiental) sol restringir les àrees de distribució de la majoria de lepidòpters, encara que també afavoreix l’expansió de les espècies oportunistes.
Per la mateixa constitució dels lepidòpters, el vent fort pot ser un factor de despoblament (raó de la manca de molts lepidòpters a illes petites). Als Països Catalans es troben força barreres i límits de l’àrea de distribució de moltes espècies de lepidòpters. Per exemple, al litoral trobem els punts més nòrdics d’espècies africanes (Charaxes jasius); i els Pirineus o el Montseny són els límits meridionals de la dispersió de moltes espècies europees (Iphiclides podalirius, Lycaena helle, etc.). Allò que per a uns lepidòpters és una barrera, per a d’altres és l’únic lloc on poden viure. Aquest és el cas de les espècies criòfiles, com Parnassius apollo, que tenen distribucions boreoalpines. A les terres catalanes tenim molts relictes glacials de les glaciacions quaternàries (paleoendemismes) que es distribueixen seguint les altes muntanyes (gènere Erebia). Aquestes espècies, que tenen poblacions aïllades geogràficament (distribució disjunta) poden tenir moltes subespècies, amb diferents èpoques d’eclosió, diferent nombre de generacions, i altres adaptacions, sense perdre el caràcter específic. Per exemple, Parnassius apollo vola a les platges de Sibèria, cosa inconcebible a Catalunya i a l’Europa central i meridional, on és una espècie d’alta muntanya. Algunes poblacions de lepidòpters es restringeixen a llocs extraordinàriament reduïts (de pocs centenars a desenes de metres), que els entomòlegs anomenen "caçadors".
Les inundacions i els anys especialment secs i calorosos determinen la desaparició de lepidòpters a determinades zones. És conegut el cas d’Anglaterra, on l’estiu de 1959 va fer que moltes espècies no s’hagin tornat a trobar. Tanmateix, veiem que la distribució d’un lepidòpter s’ha de considerar com una situació puntual d’un procés dinàmic, i no solament a escala geològica, sinó també humana. En sentit contrari, d’expansió, a Catalunya tenim un cas comprovat: el d’Araschnia levana; aquest nimfàlid travessà els Pirineus per la zona oriental, després de descobrir-se per primera vegada a la península Ibèrica a la Vall d’Aran (1962) i actualment s’estén seguint els cursos d’aigua. Però, atès el limitat coneixement biogeogràfic de la majoria de lepidòpters catalans, s’ha d’anar amb compte amb les dispersions aparents, causades pel descobriment de l’espècie a llocs on ja vivia però no s’havia trobat. Finalment, indicarem que les àrees de distribució dels lepidòpters també són afectades pels canvis periòdics de l’activitat solar, que ocasionen variacions cícliques del clima.
La importància de cada família al món, a Europa i als Països Catalans ja queda reflectida en les xifres incloses a l’apartat següent. En general, hom pot dir que els lepidòpters colonitzen pràcticament tota la terra (llevat de l’Antàrtida) i tot tipus d’ecosistemes terrestres. Al llarg dels diferents apartats ja hem exposat les diverses circumstàncies que han permès aquest èxit ecològic. Aquí tan sols farem unes consideracions de les conseqüències que això té en la distribució i dels llocs on s’adapten millor, així com de les màximes condicions que poden suportar.
Els lepidòpters s’han adaptat als boscos, les altes muntanyes (mentre no hi hagi neus perpètues), les jungles, els aiguamolls, els prats i les taigàs, les zones àrides (llevat de les àrees desèrtiques extremes), etc. Tanmateix, dins d’aquests biòtops, els lepidòpters prefereixen els climes temperats. S’han trobat lepidòpters a llocs inhabitables, sense aigua, com deserts i blocs de gel, però es tracta d’espècies migradores que s’han desviat de la ruta, o són exemplars portats pel vent. Als deserts més inòspits s’han observat nimfàlids satirins; i espècies del gènere Sphinx, així com molts microlepidòpters, s’han trobat congelats a les glaceres d’alta muntanya.
A les nostres latituds, prefereixen temperatures altes i, quan fa fred, es veuen pocs lepidòpters actius. Però als tròpics passa el contrari: l’activitat s’observa als llocs ombrívols i a les hores de menys calor. El fet que no visquin a moltes zones desèrtiques no és tant per la calor del dia, sinó pel fred del vespre. A força espècies plurivoltines catalanes s’ha comprovat que el naixement dels adults de la generació de primavera es retarda a mesura que augmenta l’altura; en canvi, a la generació d’estiu l’eclosió és simultània. Això significa que, a partir d’una determinada temperatura, ja no és favorable que faci més calor. Les poblacions més abundants de lepidòpters es donen a les zones d’altitud mitjana, vegetació molt diversa i clima sense canvis bruscos ni vents forts.
La importància de l’altitud varia segons la latitud. A les zones boreals els lepidòpters no passen dels 1000 m; a Centreuropa, molts ultrapassen els 2000 m; a la regió mediterrània els trobem fàcilment a 3000 m; als Andes arriben a 4000 i a l’Hindu Kuch als 5000, on s’alimenten dels líquens i les poques plantes superiors (gramínies) que són adaptades a aquestes altituds. S’han arribat a observar lepidòpters, concretament del grup dels satirins, volant al Tibet, a 6300 m d’altitud, i del gènere Colias a 6200 m.
Sistemàtica i filogènia
Carl von Linné, al segle XVIII, ja reuní, a la desena edició del "Systema Naturae" tots els lepidòpters dins d’un ordre, que s’ha mantingut fins a l’actualitat. Malgrat això, la classificació dels lepidòpters ha variat molt des d’aquell temps en què els lepidòpters foren agrupats tots en tres gèneres: Papilio, Sphinx i Phalaena; aquesta agrupació coincidia més o menys amb les espècies diürnes, crepusculars i nocturnes (noms donats per Latreille a començament del segle XIX). Una divisió molt popular fou la de Boisduval (1840): ropalòcers (antenes en forma de maça) i heteròcers (antenes d’altres formes), mots que avui en dia se segueixen usant. Igual passa amb els, de vegades, imprecisos noms de microlepidòpters i macrolepidòpters. A final del segle passat, Comstock definí dos grups: Jugatae i Frenatae, segons el sistema d’acoblament de les ales. Aquesta classificació coincideix amb la de Tylliard (1918) en: homoneures i heteroneures, de la qual ja hem parlat i que es basa en la nervació alar. La divisió actual més acceptada és la que figura a l’annex d’aquest volum, on s’han superposat alguns d’aquests grups antics.
Considerarem, doncs ací, l’ordre dels lepidòpters (Lepidoptera) subdividit en dos subordres, el dels zeuglòpters (Zeugloptera) i el dels glossats (Glossata), cadascun dels quals inclou diverses superfamílies. La classificació emprada pel que fa a les categories elevades ha estat elaborada segons la sistemàtica de Common (1975) i Heppner (1977), amb algunes modificacions de Kristensen (1984, 1985) adoptades per Léraut (1980), i alguns canvis recents de Razowski (1981), E. Schmidt Nielsen (1982), G. Luquet i J. Minet (1983), Heppner (1984), G. Luquet (1984-85), H. Toulgret (1985), J. Minet (1984, 1985) i Kristensen (1986). Hi ha altres tendències que varien d’aquesta i reuneixen les famílies d’una manera diferent. L’adoptada ací es fonamenta en els estudis més recents de filogènia i evolució dels lepidòpters.
La persistència de les famílies, que no han sofert tantes variacions, fa suposar que tenen una superior coincidència amb la filogènia real. En canvi, si hom baixa a les subfamílies, tribus, gèneres i subgèneres, les modificacions són encara més notables (sovint excessives) que a les categories superiors. Per aquest motiu gairebé mai no hi fem referència. En els nivells de gènere i espècie les dificultats, i àdhuc les discussions entre els especialistes, en la determinació dels exemplars poden arribar a ésser considerables, malgrat que utilitzen freqüentment la genitàlia.
Pel que fa a la filogènia dels lepidòpters, per la seva constitució fràgil i poc perdurable, no podem establir arbres filètics detallats. Malgrat això, hi ha actualment força qüestions aclarides, gràcies als estudis de morfologia comparada i als fòssils trobats, per exemple, en resines, com l’ambre. Hom creu que, procedent d’insectes més primitius, i a la frontera entre les eres Primària i Secundària, es formà un embrancament a partir del qual posteriorment es diferenciaren els tricòpters dels lepidòpters. Aquesta separació tingué lloc al Triàsic, fa uns 200 milions d’anys. Els primers insectes considerats com a lepidòpters són els micropterígids, els adults dels quals encara no han desenvolupat l’espiritrompa i mantenen les mandíbules funcionals. Aquesta i altres famílies primitives (com els eriocrànids i els hepiàlids) són homoneures, és a dir, les quatre ales amb idèntica nervació. Totes aquestes famílies que hem citat perduren actualment, i constitueixen subordres diferents; i, fins i tot, el primer s’havia considerat un ordre a part dels lepidòpters.
Progressivament, els lepidòpters varen anar adquirint els caràcters típics de l’ordre al llarg de tota l’Era Secundària, en evolució paral·lela a la de les plantes amb flors (la coevolució que ja hem referit). L’última adquisició fonamental (que els nepticúlids i els incurvàrids encara no posseeixen) és la separació dels orificis copulador i ovipositor, característica del subordre dels ditrisis (Ditrysia), el qual compta amb més del 90% de les espècies de lepidòpters. Aquestes es varen seguir modificant durant l’Era Terciària, en què s’acabaren de determinar la resta de famílies existents actualment. D’aquesta permanència dels caràcters familiars, i del fet que la taxonomia moderna no es troba tan desencaminada, en tenim una prova en la constatació que la majoria de lepidòpters fòssils trobats es poden classificar dins les famílies actuals com si fossin espècies vivents.
Els grups de lepidòpters
Els microlepidòpters primitius són els que originaren tot l’ordre, concretament els micropterígids (1 Micropterix tumbergella), els eriocrànids (2 Eriocrania semipurpurella), els diminuts nepticúlids (4 Stigmella malella), els adèlids (5 Adela reannurella) i els tisquèrids (3 Tischeria ekebladela). Dins dels ditrisians (la resta de microlepidòpters), més evolucionats, destaquen els minadors de fulles, com els gracil·làrids (10 Gracillaria syringella, la tinya de les liles). Dels tinèids, representem Nemapogon granella (6), que es coneix com a falsa tinya, cuc del blat i arna falsa del gra. Una de les famílies de morfologia larval més peculiar és la dels psíquids(8 mascle de Canephora unicolor): Dels plutèl·lids, indiquem la tinya de la col (9 Plutella xylostela), també coneguda per tinya de les crucíferes, del lligabosc i de la colza. Finalment, una família que provoca força plagues agrícoles, la dels gelèquids: representem la tinya de la remolatxa (7 Scrobipalpa ocellatella).
Servei de Fotografia/Román Montull.

Els microlepidòpters formen una agrupació artificial realitzada, a més, per entomòlegs europeus. Això fa que algunes famílies considerades de microlepidòpters tinguin representants exòtics molt més grossos que determinats macrolepidòpters d’aquí. Aquest és el cas dels ecofòrids: a la làmina hi ha una arna domèstica, Hofmannophila pseudospretella (6), que també és comensal als nius d’ocells. Els coleofòrids es distingeixen per portar sobre l’abdomen una doble renglera de petites espines (10 Coleophora hemerobiella). Els tortrícids constitueixen una de les famílies més perjudicials per a l’agricultura: un bon exemple és el cuc o corc de les pomes i peres (3 Cydia pomonella), parent de la tinya oriental (C. molesta); també ho són els rosegadors de la pell: Capua reticulana i Pandemis heparana (4). Una subfamília dels tortrícids (antigament considerada família independent) és la dels coquilins, de la qual senyalem el cuc del raïm, Eupoecilia ambiguella (1), una de les espècies que també es coneix per cuc filferro. La peculiar forma de la família dels alucítids, que no té res a veure amb les vulgarment anomenades alucites (arnes habitualment de la família dels gelèquids), la podem apreciar amb Alucita grammodactyla (9). No menys característica és la morfologia dels pterofòrids, cosa que podem comprovar observant Stenoptilia pterodactyla (7). Dues de les famílies més importants dels lepidòpters són la dels piràlids i els cràmbids, que alguns autors agrupen en una sola. A la primera pertanyen Galleria mellonella (2), conegut per arna de la cera i poll de les arnes: i Ephestia kuehniella (5), l’arna de la farina, i E. elutella. Finalment, Chilo phragmitella (8), de la segona família, és un dels barrinadors de l’arròs.
Servei de Fotografía/Román Montull.
De la vuitantena de famílies que hom suposa presents als Països Catalans, en considerarem 73 com a pertanyents, amb seguretat, a la nostra fauna. Algunes, especialment de microlepidòpters, no han estat mai citades en terres catalanes perquè no han estat estudiades, però hom pot considerar la seva presència segura. Per la manca d’estudis sobre determinats grups, de moltes d’aquestes famílies poca cosa en podem dir; generalment es tracta de microlepidòpters, espècies rares, de biologia desconeguda o que viuen a llocs amagats; tanmateix, considerant la probabilitat que el lector es pugui trobar amb les diferents espècies, hem seleccionat els tipus més representatius, que són els que descrivim en aquest apartat. Tots ells són autòctons; o sigui, no s’hi inclouen les espècies introduïdes per raons col·leccionistes o econòmiques, com és el cas de la papallona de la seda (bombícid), dels urànids, ramèids, helicònids, ofidèrids, mòrfids i altres famílies importants, però exòtiques al nostre país. L’ordenació dels grups que seguim a continuació ha estat realitzada començant pels més antics i acabant pels més evolucionats. Tanmateix cal recordar que l’evolució no és mai lineal sinó arborescent.
Els micropterigoïdeus
Els micropterígids (Micropterigidae) són papallones de mida petita i d’activitat diürna.
Les especials característiques morfològiques, úniques entre els lepidòpters, han fet que fossin considerats com un ordre a part, si bé actualment formen l’única família de la superfamília dels micropterigoïdeus (Micropterigoidea), l’única del subordre dels zeuglòpters, constituïda per un centenar d’espècies a tot el món, 15 de les quals són presents a Europa. Els adults no tenen espiritrompa, masteguen grans de pol·len amb les mandíbules funcionals. La nervació de les ales anteriors és igual que la de les posteriors. Les erugues tenen les antenes molt llargues i falses potes als vuit primers segments abdominals. La crisàlide també té trets primitius (tipus dèctic).
Els eriocranoïdeus
De la superfamília dels eriocranioïdeus (Eriocranioidea), els eriocrànids (Eriocraniidae), amb uns 50 representants a tot el món i uns 15 a Europa, són petits microlepidòpters d’activitat normalment diürna. Es tracta del segon grup més antic: tenen mandíbules, però no funcionals, i una rudimentària espiritrompa. També són homoneures (ales anteriors i posteriors amb idèntica nervació). Les femelles tenen un ovopositor llarg i perforant. Les erugues són minadores de fulles i no tenen aspecte d’eruga perquè són blanquinoses i àpodes. La crisàlide també és dèctica. La família dels eriocrànids pertany al subordre dels glossats, com totes les que comentarem a continuació, però és tan diferent de morfologia, que a més d’ésser considerada una superfamília i un grup a part (el dels dacnonifis), alguns autors en fan un subordre.
Els hepialoïdeus

De la família dels hepiàlids, l’espècie eurasiàtica Triodia sylvina (un mascle a la fotografia) és la més comuna als Països Catalans i també a la resta de la Península. L’adult vola els mesos d’agost i setembre i té una envergadura alar de 33 mm. L’eruga viu sota terra, on menja les arrels de diverses plantes ruderals (paradelles, malves, porpres, etc.).
Jordi Vidal/Col·lecció Vilarrúbia.
Els hepiàlids (Hepialidae), única família de la superfamília dels hepialoïdeus (Hepialoidea) formen, d’entre els grups menys evolucionats, el que té representants més grossos. Reuneixen aproximadament mig miler d’espècies, de les quals una vintena són presents a Europa i cinc als Països Catalans. És l’última família d’homoneures que tractarem, i alguns autors en fan un subordre, el dels exoporis (Exoporia). Són crepusculars i de coloració apagada. Les erugues són endogees, mengen arrels i poden viure anys. Són de distribució preferentment austral.
Els nepticuloïdeus
La superfamília dels nepticuloïdeus (Nepticuloidea) és la primera del grup dels mioglossats (Myoglossata) al qual pertany la resta de lepidòpters que tractarem ací. De les dues famílies que engloba, la dels nepticúlids (Nepticulidae) és la més important: conté les espècies més petites de tot l’ordre, algunes arriben a fer menys de 2 mm. Són de coloració predominantment fosca i les erugues són endòfites (minadores de fulles). Als Països Catalans tenim el gènere Stigmella. L’altra família, la dels opostègids (Opostegidae), la componen microlepidòpters, que comprenen una cinquantena d’espècies, de les quals una dotzena viuen a Europa. Són de coloració blanquinosa i únicament amb 3 o 4 nervis alars. Hi ha autors que, dins d’aquesta superfamília, també col·loquen els tisquèrids, igualment heteroneures.
Els incurvarioïdeus
Els incurvarioïdeus (Incurvarioidea), que reuneixen mig miler d’espècies, un centenar de les quals viuen a Europa, són microlepidòpters caracteritzats perquè tenen uns palps labials evidents, i, les femelles, un agut i consistent ovipositor. Les erugues viuen dins la planta nutricia i, de vegades, crisaliden dins d’embolcalls vegetals. Es un grup força heterogeni, que reuneix quatre famílies, com la dels adèlids (Adelidae), petites papallones diürnes que destaquen per tenir antenes llarguíssimes, de vegades el quàdruple del cos, i més. Alguns autors consideren aquesta superfamília com una família, i d’altres la tracten com un subordre. Són heteroneures, com totes les famílies que comentarem a continuació.
Els tisquerioïdeus
L’única família dels tisquerioïdeus (Tischerioidea), els tisquèrids (Tischeriidae), reuneix una cinquantena d’espècies de microlepidòpters molt petits i d’ales posteriors molt estretes. Tenen espiritrompa ben desenvolupada i són crepusculars o nocturns. Les erugues són minadores i la crisàlide és endòfita. Hom en coneix una cinquantena d’espècies, de les quals un centenar es distribueixen per Europa. Alguns autors consideren aquest grup com una família dels nepticuloïdeus. En qualsevol cas, són els últims monotrisians que tractarem.
Els tineoïdeus: arnes i tinyes típiques
L’important grup dels tineoïdeus (Tineoidea) és format per sis famílies de microlepidòpters; recentment ha estat revisat per Kyrki (1983), igual que els iponomeutoïdeus, si bé ací considerarem la classificació tradicional atès que en coneixem poques espècies als Països Catalans. És la primera superfamília de ditrisians, i en tractarem només les tres famílies més importants.
Els psíquids: arnes d’estoig
Els psíquids (Psychidae) són els més grossos, d’activitat nocturna, crepuscular i, de vegades, diürna. La característica principal és que els mascles tenen les escates transformades en pèls, mentre les femelles solen ser àpteres i fins i tot apodes, de la qual cosa resulta un aspecte vermiforme. Aquestes normalment no surten de l’embolcall pupal on han nascut (com una mena d’estoig), i des d’on atreuen els mascles. N’hi ha que són partenogenètiques. Les erugues també viuen dins l’embolcall que es construeixen amb trossos de la planta hoste, on crisaliden. Reuneixen un miler d’espècies, però només unes 150 viuen a Europa.
Els tinèids
Els tinèids (Tineidae), no tan petits, volen de nit. Tenen les ales de colors apagats, de vegades daurats. Les erugues mengen restes d’animals o vegetals (plomes, banyes, pell, fusta, etc.), i es donen casos de comensalisme (als nius d’ocells, mamífers i formiguers). Algunes viuen a les cases: l’arna de les estores (Trichophaga tapetzella) i les arnes de la roba i la pell (Tinea pellionella i Tineola bisselliella). Aquestes espècies domèstiques solen ésser cosmopolites. Reuneixen aproximadament 2500 espècies, de les quals unes 150 viuen a Europa.
Els gracil·làrids
Els gracil·làrids (Gracillariidae), una família d’unes 2500 espècies (de les quals unes 250 viuen a Europa), són molt petits, diürns o nocturns, amb diversos colors a les ales anteriors; les posteriors són pràcticament lineals, amb llargues prolongacions (pilositat). En posició de repòs, el cos forma un angle amb la superfície molt característic. Les erugues són minadores, com les del gènere Phyllonorycter, del qual tenim representants als Països Catalans.
Els iponomeutoïdeus: tinyes dels fruiterars i afins
Els iponomeutoïdeus (Yponomeutoidea) són microlepidòpters d’activitat nocturna. En conjunt reuneixen uns quants milers d’espècies. Hi ha erugues minadores, i d’altres gregàries, que uneixen les branques i les fulles amb fils de seda. És un grup molt heterogeni que agrupa set famílies considerades algunes d’elles com subfamílies per alguns autors. Els iponomèutids (Yponomeutidae) solen tenir ales blanquinoses puntejades de negre. La posició de repòs dels plutèl·lids (Plutellidae) és característica perquè situen les antenes erectes. Els argirèstids (Argyresthiidae) descansen col·locant-se obliquament al substrat i amb l’abdomen aixecat. Algunes espècies constitueixen plagues de la prunera (Yponomeuta padella) i d’altres arbres fruiters (Argyresthia pruniella). A la primera família que hem citat pertanyen les tinyes: el corc de capoll (Prays oleae) i P. citri que, com els seus noms indiquen, ataquen l’olivera i els cítrics. Dins una altra família —la dels acrolèpids (Acrolepiidae)— trobem la tinya del porro (Acrolepiopsis assectella).
Els gelequioïdeus: palometes del blat, palometes d’estoig, tinya de la patata i afins
La important superfamília dels gelequioïdeus (Gelechioidea) engloba una vintena de famílies de microlepidòpters, la majoria de les quals tenen representants als Països Catalans, però han estat poc o gens estudiades. Parlarem únicament de les quatre famílies més conegudes.
Els ecofòrids
Els ecofòrids (Oecophoridae) són una altra important família a escala mundial (són unes 5000 espècies, unes 200 europees). Els que viuen aquí són petits i habitualment nocturns. Tenen coloracions molt variades i palps prominents. Les erugues són fitòfagues i abasten les parts més diverses de la planta, i, fins i tot, restes vegetals i animals. Hi ha espècies comensals d’ocells i de l’home.
Els elaquístids
Els elaquístids (Elachistidae) tenen les ales estretes, les posteriors amb fines prolongacions (pilositats). Les erugues són endòfites, i, algunes, minen les fulles. En viuen unes 150 espècies a Europa.
Els coleofòrids: palometes d’estoig
Els coleofòrids (Coleophoridae) reuneixen unes 1500 espècies, de les quals aproximadament 350 viuen a Europa. Són papallones petites o molt petites, nocturnes, d’ales estretes i piloses. La posició de repòs és característica, car disposen ambdues antenes com a prolongació del cos. Les erugues construeixen galeries a les fulles i viuen dins d’un embolcall, com una beina, és diferent a cada espècie.
Els gelèquids
Els gelèquids (Gelechiidae) són papallones petites o molt petites, de coloració habitualment apagada i de vida nocturna. Les ales posteriors tenen forma trapezoïdal i l’àpex agut, a part de les fines perllongacions d’aspecte pilós. Hi ha espècies que són plagues, com la tinya de la patata (Phthorimaea operculella), el cuc rosat del cotó (Pectinophora gossypiella) i una de les palometes del blat (Sitotroga cerealella). Reuneixen unes sis mil espècies, de les quals mig miler viuen a Europa i unes 300 als Països Catalans.
Els cossoïdeus

El corc dels troncs, Cossus cossus, femella. El seu nom al·ludeix al fet que l’eruga, de les més grosses i longeves, excava galeries als salzes, als pollancres i als arbres fruiters. Hom les pot detectar gràcies als pilons d’excrements i serradures que es formen al peu de l’entrada de la galeria i que, a més, desprenen una olor característica ("l’olor de Cossus"), que també presenten alguns bolets. La substància que la produeix és l’àcid pirolignós, conegut comunament per vinagre de la fusta.
Jordi Vidal/Col·lecció Vilarrúbia.
De les dues famílies de cossoïdeus (Cossoidea), la que té més representants als Països Catalans (set espècies) és la dels còssids (Cossidae), macrolepidòpters de cos robust, poc acolorit, alguns molt grossos. Són nocturns i no tenen espiritrompa, a part d’altres caràcters primitius. El seu cicle biològic pot durar alguns anys. Les erugues són endòfites, algunes gegantines, de gran longevitat i, de vegades, provoquen una plaga, com Zeuzera pyrina (de l’avellaner), que és un dels molts insectes xilòfags que es coneix pel nom de barrinador de la fusta. Al món en viuen unes 700 espècies, i unes 10 a Europa. L’altra família, la dels limacòdids (Limacodidae) el nom de la qual es refereix a l’aspecte de les erugues, que recorda el dels llimacs, comprèn lepidòpters de grandària mitjana o petita i activitat nocturna. Alguns autors la situen dins la superfamília dels zigenoïdeus. De les 1500 espècies que existeixen al món, al nostre país són presents les tres úniques que es distribueixen per Europa.
Els tortricoïdeus: cuc verd de la vinya, cuc del raïm, cigarrers, cucs dels brots i afins
Dins la superfamília dels tortricoïdeus (Tortricoidea) l’única família és la dels tortrícids (Tortricidae), una de les més importants dels microlepidòpters (10 000 espècies al món, 1000 a Europa i 400 als Països Catalans). Són d’activitat nocturna i tenen les ales anteriors rectangulars i les posteriors trapezoïdals; en estat de repòs les pleguen de manera semblant a una teulada. Els palps sobresurten per davant. Les erugues, verdoses, elaboren un embolcall torçant (i d’aquí el nom de la família) les fulles. La seva importància econòmica és notable, car hi trobem moltes espècies que provoquen plagues, sobretot agrícoles: el cuc del raïm (Lobesia botrana), la cnefàsia (Cnephasia pumicana), el cuc revoltó o cuc verd de la vinya (Sparganothis pilleriana), el tòrtrix dels alzinars (Tortrix viridana), les cuques del clavell (Cacoecimorpha pronubana i Epichoristodes acerbella) i alguns dels anomenats cigarrers: Archips xylosteana i A. rosana, aquest últim també conegut per cuc dels brots.
D’entre les diverses subfamílies en què hom divideix els tortrícids, en citarem la més diferenciada: la dels coquilins (Cochylinae), que alguns autors consideren una família independent. Comprèn més de mig miler d’espècies (prop de 300 a Europa i 50 als Països Catalans), algunes de les quals són perilloses per a l’agricultura.
Els sesioïdeus: borinots vespa i afins
De les tres famílies que engloba la superfamília dels sesioïdeus (Sesioidea), parlarem només de la més important, la dels sèsids (Sesiidae), formada per petits macrolepidòpters diürns que es distingeixen de la resta de famílies pel seu aspecte d’abelles, vespes o himenòpters en general. Aquest mimetisme batesià l’han aconseguit per la pèrdua de les escates de les ales posteriors (que queden translúcides) i parcialment de les anteriors; per la possessió de ratlles negres, grogues i vermelles a l’abdomen; i per la realització del vol de manera semblant als himenòpters. Les erugues són endòfites o, de vegades, endogees. És una família important a Amèrica del Sud. Reuneix unes 1500 espècies, de les quals 80 són europees i unes 40 són representades al nostre país.
Els alucitoïdeus
De la superfamília dels alucitoïdeus (Alucitoidea), l’única família dels alucítids (Alucitidae), també anomenats orneòdids (Orneodidae), reuneix unes 150 espècies (15 europees). Són papallones petites i fràgils, de vida nocturna, que es diferencien clarament perquè tenen les ales partides en sis segments transversals, que recorden la forma de les plomes. Les erugues són endòfites i antòfagues.
Els pteroforoïdeus
Els pteroforoïdeus (Pterophoroidea), que comprenen només la família dels pterofòrids (Pterophoridae), una família amb unes 1000 espècies, de les quals 200 són presents a Europa, són petites papallones que volen de nit i tenen un aspecte inconfusible. Això els ve del fet de tenir les ales partides, formant 2, 3 o 4 lòbuls, i perquè, en repòs, el cos i les ales adopten la forma d’una T. Hi ha erugues endòfites i d’altres que s’emboliquen teixint les fulles amb la seda.
Els piraloïdeus: arnes del gra i afins
La superfamília dels piraloïdeus (Pyraloidea), comprèn la família dels piràlids (Pyralidae), que reuneix unes 15 000 espècies (unes 300 a Europa) i la dels cràmbids (Crambidae), amb unes 20 000 espècies (unes 350 a Europa).
Els piràlids
Els piràlids constitueixen una de les principals famílies dels microlepidòpters, però amb poques espècies a casa nostra. A altres continents, hi ha representants tan grossos que ja es poden considerar macrolepidòpters; de fet, és un grup intermedi entre aquests i els microlepidòpters. Aquestes papallones nocturnes han desenvolupat un autèntic timpà a l’abdomen. En posició de repòs, col·loquen les ales planes sobre el cos. Les erugues, que poden constituir plagues, mengen gran diversitat de materials, i algunes són domèstiques; són conegudes com a arnes, com la del gra (Pyralis farinalis) i la del farratge (Hypsopigia costalis).
Els cràmbids
Els cràmbids formen la família amb un nombre més gran d’espècies de tots els microlepidòpters, d’entre els quals, a més, són dels més grossos. Malgrat això, als Països Catalans no assoleixen la importància ecològica que tenen a altres zones. Es tracta d’un grup molt semblant a l’anterior, i amb preferències nocturnes. Els palps sobresurten molt del cap i això els dona un aspecte diferenciador de les altres famílies. Freqüentment, les ales són de color de palla i, en repòs, envolten l’abdomen. Aleshores, en disposar-se en posició vertical a sobre les gramínies (habitualment les plantes hoste) aconsegueixen una bona cripsi. Algunes erugues causen flagells a l’agricultura, com Ostrinia nubilalis, un dels barrinadors del blat de moro.
Els zigenoïdeus: zigenes i afins
Dels zigenoïdeus (Zygaenoidea) només ens podem referir als zigènids (Zygaenidae), una família que reuneix unes 1200 espècies, de les quals 60 són a Europa i unes 32 als Països Catalans. Són tots diürns, de dimensions petites o mitjanes i colors aposemàtics metàl·lics, premonitoris de la seva toxicitat, la qual conserven en totes les seves fases. Els de la subfamilia dels procridins (Procridinae) són de color verd marronós mentre que els zigenins (Zygaeninae) són blaus, vermells i blancs, combinats en coloracions patró. Aquest mimetisme mullerià els fa inconfusibles. Les erugues són rabassudes, amb el cap retràctil, i la crisàlide allargada i fusiforme.
Els hesperioïdeus
La família dels hespèrids (Hesperiidae), l’única de la superfamília dels hesperioïdeus (Hesperioidea), inclou papallones de dimensions mitjanes o petites i ales curtes i arrodonides. Volen de dia amb rapidesa i zigzaguejant. Tant els adults com les erugues tenen el cap gros en proporció al cos. Les antenes són separades a la base i acabades en forma de ganxet. És un important grup cosmopolita, especialment nombrós a Amèrica del Sud; de les 5000 espècies que comprèn, hom en coneix una cinquantena a Europa i 25 als Països Catalans.
Els papilionoïdeus
La superfamília dels papilionoïdeus (Papilionoidea) engloba la gran majoria de les espècies diürnes típiques. Són de colors molt vistents i d’aspecte molt variable, tot i que tenen en comú la forma de les antenes, que acaben en maça, i la posició de les ales en repòs, que es disposen en vertical. Aquests caràcters els comparteixen amb els hesperoïdeus, també lepidòpters diürns.
Els papiliònids: papallones rei i afins
Els papiliònids (Papilionidae) són grossos macrolepidòpters diürns, amb ales de colors vius. L’aspecte extern és molt variable i amb dissenys atractius. Algunes espècies són en perill d’extinció per l’abusiva captura dels col·leccionistes. Hi ha representants tropicals mimètics amb famílies tòxiques, com els helicònids i nimfàlids danaïns (Danainae). Les erugues tenen una glàndula postencefàlica evaginable. Malgrat l’escassa representació als Països Catalans (6 espècies), és una família àmpliament distribuïda a la zona tropical. Hom en coneix un total de 700 espècies, de les quals únicament se’n coneixen 12 a Europa.
Com a exemples de papiliònids, una de les famílies de papallones més vistents del nostre país, podem esmentar la papallona apollo (Parnassius apollo). Aquesta i l’espècie congenèrica P. menosyne són boreoalpines, amb poblacions aïllades als diversos massissos d’Europa; aquest fet i l’afany dels col·leccionistes ha fet que, de la primera, se n’arribessin a descriure prop de 300 subespècies, la majoria de les quals són infonamentades. És una espècie en perill d’extinció a alguns països, on la llei la protegeix (no així al nostre). D'altra banda, la papallona rei o rei de papallones (Papilio machaon), és una espècie estesa per tota la Península, igual que la papallona zebrada Iphiclides feisthamelii, pròxima i també molt vistent; hi ha una altra papallona zebrada, Iphiclides podalirius, però només vola a la Vall d’Aran i més al nord, a Europa.
Els pièrids: blanquetes, groguetes i afins
Els pièrids (Pieridae), una família que reuneix 1600 espècies aproximadament, és una mica més ben representada a Europa (45 espècies) i als Països Catalans (21 espècies) que no l’anterior. Són macrolepidòpters de grandària mitjana que volen de dia, sovint lentament. Tenen antenes acabades en forma de martell. El color dominant de les ales és el blanc groguenc amb taques negres. Freqüentment presenten polimorfisme sexual i estacional, i algunes (exòtiques) mimetisme amb espècies tòxiques. Les erugues poden ser perilloses per a l’agricultura (cosa inhabitual entre els ropalòcers) atès que mengen crucíferes i lleguminoses. El cas més conegut és el de l’anomenat cuc de la col, que no és més que l’eruga de la papallona blanca de la col (Pieris brassicae).
Com a exemples de pièrids podem citar Colias crocea, una espècie abundant arreu, que presenta diverses formes de femelles (polimorfisme sexual). I també Gonopteryx cleopatra, espècie comuna i coneguda amb el nom de llimonera, que també s’aplica, però, a la congenèrica G. rhamni. D’aquesta mateixa familia trobem als Països Catalans alguns representants del gènere Pieris, del qual, a part la papallona de la col (Pieris brassicae), podem citar P. rapae i P. napi. Del gènere Anthocharis, hi ha A. cardamines i A. euphenoides. També abunden Pontia daplidice, Aporia crataegi i Euchloe ausonia. Com a característiques generals d’aquesta família podem dir que els ous tenen forma de petits barrils allargats amb estries, que les erugues són molt cilíndriques, llises i llargues, i algunes mirmecòfiles, i que algunes crisàlides es fixen amb la part cefàlica enlaire, assegurades per un cinturó de seda.
Els nimfàlids: vanesses, dameres, paons de dia i afins
Els nimfàlids (Nymphalidae) constitueixen una família molt important de macrolepidòpters diürns, de gran popularitat i dimensions mitjanes o grans, sovint amb dissenys alars força acolorits (excepte la subfamilia dels satirins), raó per la qual també són apreciades pels col·leccionistes. Les subfamílies són ben diferenciades, i fins fa poc es consideraven famílies. Una característica notable —no compartida pels libiteïns— és que només usen el segon i el tercer parell de potes, car el primer el tenen atrofiat, amb major o menor grau. Reuneix un bon nombre d’espècies (unes 7000), de les quals 200 són presents a Europa i 87 són conegudes al nostre país.
La subfamilia dels nimfalins (Nymphalinae) és la que té colors més llampants, de vegades amb cues i reflexos metàl·lics a les ales. Hi ha espècies hivernants i migradores. Les erugues també són espectaculars i de colors aposemàtics (perquè són tòxiques). Les erugues dels gèneres Argynnis, Vanessa, Inachis i altres, tenen pèls punxents, que poden ser endurits i ramificats. Als Països Catalans en tenim 40 espècies.
Un dels nimfalins més abundants i estès per tota la Península és la papallona reina (Vanessa atalanta), una espècie migradora que a les nostres latituds sol efectuar llargs desplaçaments (de centenars de quilòmetres) cap al nord; igualment fan, entre altres, la vanessa dels cards (Cynthia cardui), i la de les ortigues (Aglais urticae). El paó de dia (Inachis io) un dels nimfàlids més comuns a la meitat nord de la Península, vola principalment a l’estiu i els adults hivernen i no s’aparellen fins a la primavera següent, cosa que és poc habitual entre els lepidòpters. Altres nimfalins pròxims a aquests i abundants són: la papallona de l’arboç (Charaxes jasius), la papallona tornassolada (Apatura ilia), Limenitis Camilla i L. reducta, Nymphalis antiopa (negre amb una ampla vora groga), la papallona retallada (Polygonia c-album) i Araschnia levana, espècie aquesta que representa el nimfàlid més petit i, probablement, la papallona que mostra un polimorfisme estacional més accentuat.
Moltes espècies de nimfalins tenen un predomini del color taronja a l’anvers de les ales i moltes cel les geomètriques petites de diferents colors al revers, i per això i hom les anomena damers. Aquest és el cas de les papallones dels gèneres Brenthis, Boloria, Clossiana, Melitaea, Mellicta, Eurodryas i altres. En són exemples, Issoria lathonia, inconfusible per les cel les platejades del revers de les ales, que vola abundantment en dues generacions: una als mesos d’abril i maig i una altra als de juny i juliol; l’eruga és hoste de les violàcies; Eurodryas aurinia, que viu des del nivell del mar fins a més de 3000 m, i és bivoltina: la primera generació emergeix pel maig i la segona a l’agost; les erugues són molt polífagues i gregàries; i Argynnis paphia, un dels nimfalins més grossos, que abunda a la meitat nord de la Península, a una altura mitjana; la femella efectua la posta al tronc d’un arbre i les erugues mengen la closca de l’ou i hivernen dins l’escorça, fins a la primavera següent, estació en què les pròpies erugues surten a cercar les plantes nutrícies (rosàcies i violàcies). Altres nimfalins abundants són Clossiana dia, Melitaea didyma, Mellicta deione i M. parthenoides.
Els satirins (Satyrinae) tenen una gran resistència a ambients extrems. El gènere Erebia, per exemple, és dels que arriba als cims més alts. El revers de les ales posteriors és molt críptic. Les erugues, que sovint hivernen a la base de la planta hoste (habitualment gramínies), es diferencien per la protuberància bífida que tenen al final de l’abdomen. Als Països Catalans en volen 45 espècies. Amb els colors apagats que caracteritzen la família podem citar l’espècie Brintesia circe, una papallona que vola a l’estiu, preferentment en indrets secs, i s’està llargues estones a l’escorça dels arbres, on, amb les ales plegades, esdevé totalment críptica; l’eruga, com la majoria de les del seu grup, és hivernant i s’alimenta de gramínies. A la mateixa subfamília dels satirins pertanyen les melanàrgies, nom sota el qual es coneixen cinc espècies catalanes, entre elles, Meanargia lachesis, M. occitanica i M. galathea, aquesta de zones més altes i de morfologia pràcticament igual que la primera. Del mateix grup, destaquem les espècies catalanes dels gèneres Erebia, (E. meolans, E. triaria, E. neoridas i E. cassioides) i Hipparchia (H. alcyone, H. semale, H. statilinus i H. fidia). Un altre satirí abundant a tots els Països Catalans és Pyronia cecilia, que veiem a la fotografia, el qual vola de juny a agost per les zones de vegetació baixa, com la majoria de les altres papallones de la seva subfamilia. Altres espècies relacionades i abundants són P. tithonus i P. bathseba, Aphantopus hyperantus, que viu a la franja septentrional ibèrica i prefereix les zones no massa caloroses, Chazara briseis, abundant a tota la Península tret de la part de ponent, Maniola jurtina, freqüent arreu, igual que Pararge aegeria, Lasiommata megera i la diminuta Coenonympha pamphilus.
Dels danaïns (Danainae), que són tropicals, només trobem una espècie, migradora (Danaus chrysippus), que ha vingut de l’Àfrica, i que només ha estat citada recentment. També hi ha alguna citació esporàdica de D. plexippus. Un altre satirí abundant a tots els Països Catalans és Pyronia cecilia, que veiem a la fotografia, el qual vola de juny a agost per les zones de vegetació baixa, com la majoria de les altres papallones de la seva subfamilia. Altres espècies relacionades i abundants són P. tithonus i P. bathseba, Aphantopus hyperantus, que viu a la franja septentrional ibèrica i prefereix les zones no massa caloroses, Chazara briseis, abundant a tota la Península tret de la part de ponent, Maniola jurtina, freqüent arreu, igual que Pararge aegeria, Lasiommata megera i la diminuta Coenonympha pamphilus.
La subfamilia dels libiteïns (Libytheinae) també és tropical, però en tenim un representant autòcton: la papallona del lledoner (Libythea celtis).
Els licènids: blavetes, coures i afins

Tres espècies comunes de licènids: a dalt, mascle i femella de Pseudophilotes panoptes, que representa un dels casos en què ambdós sexes són similars, en aquesta ocasió, de color blau; al mig, Callophrys rubi, anvers i el revers, inconfusible pel color verd de les ales; i a baix, mascle i femella de Glaucopsyche melanops.
Jordi Vidal/Col·lecció Vilarrúbia.
Els licènids (Lycaenidae), coneguts com a blavetes, són petits macrolepidòpters diürns (gairebé tan petits com els hespèrids) que constitueixen una de les famílies amb més importància ecològica. Presenten polimorfisme estacional, però sobretot dimorfisme sexual molt notable. Una situació normal és que els mascles siguin blaus i les femelles marrons (d’ací el nom de blavetes, ja que els mascles són més abundants). Hi ha espècies amb colònies molt localitzades, i d’altres que es troben entre els lepidòpters més estesos i abundants. Els mascles es concentren als tolls d’aigua i volen formant grans estols. Els ous i erugues dels licènids són molt típics: els primers perquè tenen forma discoidal, i les segones, inconfusibles, perquè no tenen l’aspecte eruciforme i semblen un porquet de sant Antoni (isòpode), car són rabassudes, curtes, aplanades i el cap no es distingeix; algunes són carnívores depredadores i d’altres mirmecòfiles.
De les aproximadament 9000 espècies de licènids existents, hom en coneix unes 110 a Europa i una seixantena al nostre país. De fet, és una família que es troba en plena efervescència evolutiva, car s’han trobat molts híbrids, clines de variació, superspècies, subespècies, espècies dubtoses i tàxons difícils de diferenciar. A les nombroses incerteses sistemàtiques, últimament s’hi ha afegit la d’incloure dins d’aquesta família els nemeòbids, antic nom dels riodínids, ara riodinins (Riodininae); d’aquesta subfamilia tropical, als Països Catalans, només n’arriba una espècie: Hamearis lucina, únic representant europeu.
Un dels licènids més petits és Pseudophilotes panoptes, de 20 mm d’envergadura alar, que representa un dels casos en què mascle i femella són similars (en aquesta ocasió, de color blau, en d’altres espècies, marró). Callophrys rubi té 28 mm d’envergadura alar i és inconfusible pel color verd de les ales, color molt poc comú no solament entre els licènids sinó en tots els lepidòpters, per la qual cosa els convé el nom de verdes (també C. avis, molt més rara); volen durant la primavera, i als indrets calorosos hi ha una segona generació d’estiu. Glaucopsyche melanops (a baix), té una envergadura alar de 24 mm, i les seves ales són menys blaves i més marronoses en les femelles; l’eruga s’alimenta de lleguminoses. La seva congenèrica G. alexis és mirmecòfila. Amb un marcat dimorfisme sexual Lysandra hispana, de 34 mm d’envergadura alar, és molt semblant a L. coridon; alguns autors situen ambdues espècies al gènere Plebejus. Algunes espècies afins a aquestes i mirmecòfiles són Maculinea alcon i M. arion. Quan, després de la tercera muda, s’activen les glàndules de Newcomer, diverses formigues— per exemple del gènere Myrmica— transporten les erugues dins del formiguer, on són alimentades i hivernen. La majoria dels licènids tenen nombrosos ocel·les dibuixats al revers de les ales. Altres licènids molt abundants són Lycaena phlaeas, una de les més esteses; L. virgaureae; Tomares ballus; Strymonidia spini, mirmecòfila; Cupido minimus, molt diminuta, com el seu nom indica; Polymmatus icarus, que forma estols de centenars i fins de milers d’exemplars; i Lampides boeticus, la qual, dotada d’unes petites cues que en repòs prenen l’aparença d’antenes, confon el predador, el qual en atacar el cap s’endú només trossos d’ales.
Els axioïdeus
Els axioïdeus (Axioidea) són papallones petites i fràgils, que constitueixen un grup petit (reuneix cinc espècies únicament) i endèmic de la regió mediterrània. Comprèn dos gèneres, amb tres espècies europees i una sola que forma part de la nostra fauna. Són semblants als drepànids i als geomètrids, i alguns autors, per això, consideren la seva única família, la dels àxids (Axiidae) dins la superfamília dels geometroïdeus.
Els drepanoïdeus
La superfamília dels drepanoïdeus (Drepanoidea) comprèn un total de 800 espècies, de les quals 25 es distribueixen per Europa i 14 formen part de la nostra fauna. Els drepànids (Drepanidae), l’única família del grup, són macrolepidòpters de mida variable, més aviat petita en el cas de les espècies que formen part de la nostra fauna, i d’activitat nocturna. Alguns autors els consideren inclosos dins la superfamília dels geometroïdeus. Hom en considera dues subfamílies ben diferents.
D’una banda, els drepanins (Drepaninae), característics pel fet de tenir els palps maxil·lars atrofiats i també, en un grau més o menys accentuat, l’espiritrompa. El seu aspecte extern recorda el dels geomètrids, però una característica inconfusible, per a algunes espècies, és la forma de falç amb què acaben les ales anteriors, que dona nom a la família (drepanon, falç). Les erugues són molt característiques; els manca el cinquè parell de falses potes i l’abdomen acaba enlairat i en forma cònica. Crisaliden fent un embolcall pupal cobert de fulles. Als Països Catalans en coneixem 6 espècies. Drepana uncinula, que presenta de 24 a 35 mm d’envergadura alar, és la més comuna a la península Ibèrica, però no a la resta d’Europa. Una altra espècie molt abundant és precisament l’única ibèrica que no té la forma de falç, Cilix glaucata, i que, a més de ser el representant més petit del grup, té la particularitat de simular, en repòs sobre una pedra, un excrement d’ocell, per tal de passar desapercebut.
D’altra banda, la subfamilia dels tiatirins (Thyatirinae), que alguns autors consideren com una família, amb el nom de cimatofòrids (Cyrnatophoridae), engloba una quarta part de les espècies, de les quals 15 viuen a Europa i 8 als Països Catalans. Els distingeix el fet de tenir l’espiritrompa i els palps ben desenvolupats i el seu aspecte general de noctúid. Els tiatirins més nombrosos a la península Ibèrica són Thyatira batis, una espècie amb 36 mm d’envergadura alar que presenta dues generacions (al maig-juny i a l’agost-setembre), i Habrosyne pyritoides, univoltina. Totes dues es troben arreu, llevat de l’alta muntanya; les erugues mengen esbarzers. Un altre representant que pot ser abundant és Tethea ocularis, bivoltina, que trobem en ambients humits; les erugues s’alimenten de pollancres, salzes i bedolls, entre altres arbres.
Els geometroïdeus: geòmetres o acanadores
De la superfamília dels geometroïdeus (Geometroidea), els geomètrids (Geometridae), actualment considerats com l’única família del grup, reuneixen macrolepidòpters de grandària variable, gairebé tots nocturns o crepusculars: És una família cosmopolita i de les més importants dins l’ordre. Tenen ales grosses en proporció al cos, fràgils i de colors clars amb molts dibuixos. Així aconsegueixen una cripsi notable sobre el sòl, que afavoreixen en disposar-hi les ales ben aplanades. A l’abdomen tenen un autèntic timpà. Algunes femelles són braquípteres, micròpteres i, fins i tot, àpteres. L’eruga encara es camufla més bé, immobilitzant-se i simulant les branques. Si les localitzem, ens resultaran inconfusibles, car només tenen dos parells de falses potes, i es mouen collocant-les directament al costat del cap. Amb aquest moviment semblen prendre mides i és molt característic, donant nom a la família (metro, amidar, geo, terra). Vulgarment se les coneix com a apamadores o agrimensores. Comprenen un total de 20 000 espècies, de les quals 1100 es distribueixen per Europa i aproximadament 500 als Països Catalans.
Un dels geomètrids més estesos i coneguts és Abraxas grossulariata, que es troba a tota la regió paleàrtica i és univoltí; la seva eruga s’alimenta d’arbres fruiters. L’espècie congenèrica A. sylvata, estesa pels boscos paleàrtics caducifolis, vola de maig a agost.
Els geomètrids presenten una gran diversitat de formes, però generalment tenen les ales febles, molt amples en relació al cos, i força dibuixades amb punts i línies trencades. Les erugues resulten inconfusibles per la manca de pseudopodis (figura 347) i la manera de moure’s que se’n deriva. Entre els geomètrids més comuns als Països Catalans podem citar Pseudoterpna coronillana, Timandra griseata, Scopula ornata i S. rubiginata (aquestes dues de llocs secs i amb dues o tres generacions superposades), Rhodostrophia sacraria, Cidaria fulvata (la seva eruga viu als rosers dels jardins i l’adult vola en una generació d’estiu), Bichroma famula, Pachycnemia hippocastanaria, Pseudopanthera macularia (de morfologia variable), Aspitates gilvaria i A. ochrearia (termófiles), Bupalus piniaria (amb un marcat dimorfisme sexual), Emerophila abruptaria, Eurranthis plummistaria (que vola de dia), i Biston hirtaria.
Els bombicoïdeus
Hom reuneix dins la superfamília dels bombicoïdeus (Bombycoidea) macrolepidòpters generalment vistents, d’activitat habitualment nocturna, pertanyents a diverses famílies, de les quals tractarem les dels lasiocàmpids (Lasiocampidae), dels esfíngids (Sphingidae) i dels satúrnids (Saturniidae).
Els lasiocàmpids
Els lasiocàmpids (2200 espècies a tot el món, amb 50 representants a Europa i 25 al nostre país) són macrolepidòpters de mida normalment superior a la mitjana. Volen de nit, excepte els mascles d’algunes espècies, com Lasiocampa quercus. Tenen el cos rodanxó i pelut, les ales amples i de colors apagats, l’espiritrompa atrofiada, i les antenes plomoses. Deixen postes agrupades i nombroses, i les erugues són de creixement lent, cobertes d’una abundant pilositat, de vegades urticant. El nom de la família ve de campé, eruga, i lasios, peluda. Algunes són gregàries i viuen a nius de seda, la qual també usen per a fabricar l’embolcall pupal. És una família ben coneguda, important sobretot als tròpics i a l’Amèrica del Sud. Aquí, alguna espècie pot provocar plaga als arbres.
Els esfíngids: borinots i esfinxs
Els esfíngids reuneixen prop de 1500 espècies especialment abundants a la zona tropical, on es troben espècies minúscules, d’ales de 4 mm d’envergadura. Hi ha abundants casos d’hibridisme. A Europa hom en coneix una trentena d’espècies i una vintena al nostre país. Són de grandària i activitat molt variables (d’1 a 20 cm d’envergadura alar), però les del nostre país són més aviat grosses i crepusculars, encara que n’hi ha de nocturnes i de diürnes. De cos ferm i fusiforme, tenen ales potents, les anteriors llargues i estretes però robustes, cosa que les fa molt aerodinàmiques. Efectivament, tenen un vol ràpid i són grans migradores. Per això és possible que visiti els Països Catalans alguna espècie no inclosa dins la vintena indicada. Tenen un òrgan acústic als palps i l’espiritrompa molt llarga (fins a 25 cm en les espècies més grosses). Alguns autors consideren aquest grup com una superfamília independent. Destaquen l’esfinx ocellat (Smerinthus ocellata i l’esfinx o papallona de la mort (Acherontia atropos, aquesta darrera també coneguda amb els noms de borinot de la calavera i cap de mort. Altres esfíngids coneguts són l’esfinx de les lletereses (Hyles euphorbiae), l’eruga de la qual és anomenada pedraferit de les lletereses, i l’esfinx de la troana (Sphínx ligustri). Entre els esfíngids més petits trobem l’espècie Hemaris fuciformis, que com H. tityus (molt semblant) protagonitza un cas de mimetisme batesià juntament amb els himenòpters: a les ales sense escates i els colors abdominals d’avís s’afegeix el vol vibrat estacionari, que els dona una gran semblança amb el sèsids. I també, l'anomenat bufaforats (Macroglossum stellatarum). Molts esfíngids realitzen llargues migracions i travessen tota la Mediterrània, o tota Europa, de cap a cap, especialment vers el N.
Les erugues dels esfíngids, que popularment hom coneix com a pedraferits, són inconfusibles per l’apèndix anal en forma de corn. Són glabres i amb ratlles transversals i colors evidents, aspecte que es reforça en posició de repòs: el cap i el tòrax aixecat, recordant l’esfinx de la mitologia, que dona nom a la família. N’hi ha que són perilloses per a l’agricultura. La crisalidació pot efectuar-se sota terra.
Aquestes conegudes papallones han rebut nombrosos noms vulgars. Seguidament assenyalarem algunes de les espècies que podem veure al nostre país: l’esfinx de les corretjoles (Agrius convulvuli), l’esfinx del pi (Hyloicus pinastri), la dels til·lers (Mimas tiliae), la del pollancre (Laothoe populi), la del roure (Marumba quercus), i la dels baladres (Daphnis nerii).
Els satúrnids: graèlsia, bruixa, cuc de seda de l’alzina, paons de nit i afins
Els satúrnids, també coneguts com a atàcids (Attacidae) són papallones grosses o molt grosses, d’activitat predominantment nocturna. Tenen ales de gran superfície i fermesa, un gran ocel·le molt típic i constant i, de vegades, cues, les antenes pectinades (en els mascles, quatripectinades, per a captar millor l’olor de les feromones) i l’aparell bucal atrofiat. El dimorfisme sexual és accentuat. Hom n’ha descrit força híbrids. Les erugues són molt vistents per les seves grandària i coloració; tenen verrugues, tubercles espinosos i pèls complicats (urticants o estel·liformes), i les seves glàndules salivals són de les més llargues dels lepidòpters. La crisàlide és protegida per un embolcall pupal molt rígid.
Entre els satúrnids figuren les espècies de papallones més grosses quant a superfície alar. El gran paó de nit (Saturnia pyri), també anomenat bruixa, és l’insecte més gros d’Europa. El petit paó de nit (Pavonia pavonia) té un marcat dimorfisme sexual, i el mascle vola de dia, contra el vent i seguint el gradient d’olor de la feromona emesa per la femella, que resta amagada. L’altra espècie catalana de satúrnid és Aglia tau (el més rar al nostre país), el mascle del qual podem trobar a les fagedes també volant de dia. La relació superfície/pes total dels satúrnids és proporcionada pel cos molsut i pesant.
La graèlsia (Graellsia isabelae) és considerada un dels lepidòpters més bells del món, cosa que, de fet, no ha estat gaire positiva per a la pròpia espècie: la persecució dels col·leccionistes nacionals i estrangers ha fet que desaparegués de diversos indrets (actualment, tot i que és protegida per la llei, a França ha esdevingut molt rara). Als Països Catalans hi és força estesa gràcies al fet que l’eruga s’alimenta de pi roig fonamentalment (i, en un grau menor, també de pinassa), que forma boscos a llocs més aviat apartats de la màxima degradació.
Els satúrnids tenen erugues grosses, molsudes, de colors vius i apèndixs pilosos. L'eruga del petit paó (Pavonia pavonia) i la del gran paó de nit (Saturnia pyri) s'assemblen molt, però aquesta darrera té els estigmes blancs en lloc de vermells. Ambdues erugues són molt polífagues, i s’alimenten de molts arbres fruiters. Els adults són univoltins; el del petit paó, més abundant que el gran paó de nit, es troba durant la primavera i, el d’aquest últim, durant els mesos de maig i juny, només a les nits.
A la zona tropical és una família important i s’hi troben espècies de les més grosses entre els lepidòpters. Als Països Catalans, on la representació és testimonial (4 espècies de les 6 europees, comparat amb les 1600 en total), tenim dues espècies introduïdes per a l’aprofitament de la seda: Samia cynthia i la papallona del cuc de seda de l’alzina (Antheraea pernyi), que es poden trobar esporàdicament a Barcelona (la segona també a les Gimnèsies), escapades dels criadors.
Els noctuoïdeus
La superfamília dels noctuoïdeus (Noctuoidea) és la més important numèricament, i la més diversificada ecològicament, de tots els lepidòpters, només comparable a la dels piraloïdeus. A més, els noctuoïdeus tenen una gran repercussió econòmica, ja que inclouen la majoria de les espècies que constitueixen flagells agrícoles i forestals importants, com les processionàries, els corcs dels horts, etc.
Els notodòntids: cues forcades, processionàries i afins
Els notodòntids (Notodontidae) reuneixen un total de 3000 espècies amb una representació modesta a Europa (55 espècies) i al nostre país, on se’n coneixen 36. Són macrolepidòpters de grandària mitjana, coloració críptica i activitat nocturna. Tenen l’espiritrompa atrofiada, el cos gros i amb abundant pilositat, la qual, sobre el tòrax, forma una cresta característica que dona nom a la família (notos, espatlla, odons, dent). En repòs, pleguen les ales en forma de teulada, sobresortint més la cresta. Grup clarament dividit en dues subfamílies que molts autors segueixen considerant famílies diferents: els notodòntids típics i els taumetopèids.
La subfamília dels notodontins (Notodontinae) inclou més del 95 % de les espècies. A Europa en viuen prop de mig centenar, de les quals 33 les trobem als Països Catalans. És un grup important a Amèrica del Sud. Es caracteritza perquè les erugues, de colors llampants, adopten postures espaoridores molt típiques, amb els dos extrems aixecats i formant una gepa. A l’extrem anal, en comptes de ventoses o pseudopodis, tenen un apèndix bífid, del qual surten uns tubs evaginables per a la defensa.

Els taumetopeïns són les conegudes processionàries. A les fotografies veiem l’adult (mascle i femella, d’envergadura alar de 35 i 44 mm, respectivament) i l’eruga de la processionària més perillosa, la del pi (Thaumetopoea pityocampa). L’adult emergeix de mitjan juny al final de juliol, i vola fins al setembre, rarament l’octubre. Les erugues, gregàries des que neixen, construeixen un segon niu ben protegit, dins del qual passen l’hivern. Crisaliden a la primavera, i la pupa pot durar de pocs mesos a sis anys. Aquestes crisàlides són les responsables de les sobtades reactivacions de plagues aparentment controlades. La processionària del roure, del mateix grup, és T. processionea.
Jordi Vidal i Ramon Torres.
Els taumetopeïns (Thaumetopoeinae) tenen la cresta esclerificada i característica de cada espècie. Les femelles tenen una bossa anal. Les erugues són les conegudes processionàries. Són urticants i gregàries: viuen dins de nius de seda i crisaliden enterrant-se en grups. Constitueixen greus plagues de les pinedes, rouredes, etc. El nombre d’espècies conegudes d’aquesta subfamilia no arriba al centenar; cap d’elles no viu a Amèrica. A Europa en trobem mitja dotzena, de les quals només tres volen als Països Catalans.
Els limàntrids: cuques peludes del suro i afins
Els limàntrids (Lymantriidae) constitueixen una família d’unes 3000 espècies, representada a Europa per 25 espècies i al nostre país per només 17 espècies. Es tracta de petits macrolepidòpters nocturns, però amb alguns mascles que volen de dia. Aquests tenen les antenes curtes i enterament bipectinades, són petits i de vol ràpid zigzaguejant. Les femelles són més grosses, amb l’abdomen molsut, dissenys alars diferents, i, fins i tot, algunes són àpteres. Tant l’adult com l’eruga presenten abundant pilositat. Tenen òrgans timpànics al tòrax i l’espiritrompa atrofiada.

Mascle i femella de Lymantria dispar, que duu aquest nom perquè mascle i femella (36 i 52 mm d’envergadura alar) no s’assemblen gens; la larva rep el nom d’eruga peluda del suro.
Jordi Vidal/Col·lecció Vilarrúbia
La paraula limàntrids significa perjudicials, amb relació al fet de contenir la família algunes espècies capaces d’experimentar sobtades explosions demogràfiques i destruir boscos sencers. De tots els lepidòpters, el gènere més devastador ha estat i és, no solament a les nostres latituds, Lymantria. L. dispar, duu aquest nom perquè mascle i femella (36 i 52 mm d’envergadura alar) no s’assemblen gens; la larva rep el nom d’eruga peluda del suro. Val a dir que aquesta i la majoria de plagues són conseqüència d’una acció humana que ocasiona una alteració ambiental, la qual provoca la desaparició o disminució d’algun factor que controla la població del lepidòpter. Altres limàntrids, no són tan perillosos com a causants de plagues. Orgyia antiqua viu a la meitat nord de la península Ibèrica, i només excepcionalment provoca plagues molt limitades. La femella, com les de les altres espècies del seu mateix gènere, és totalment àptera. Quan arriba el mascle, que vola de dia seguint el rastre de la feromona femenina, s’aparellen sense que aquesta surti de l’embolcall pupal; només en surt per a fer la posta, i encara, sobre el mateix envà pupal. Hi ha una raça partenogenètica en la qual la femella efectua la posta sense haver-se acoblat prèviament i dona lloc exclusivament a femelles. Les erugues (les femenines més grosses que les masculines) són polífagues i de coloració aposemàtica, formada per vistents pinzells de pèls de colors.
Les erugues solen ésser gregàries des de la posta (recoberta amb una barreja protectora de pèls i secreció adherent). Són urticants i presenten pinzells de pèls de colors llampants. Algunes construeixen nius i moltes causen gravíssimes plagues agrícoles i forestals, particularitat que dona nom a la família (limanterios, perjudicial). És un grup d’extraordinària incidència ecològica.
Els àrctids
Els àrctids (Arctiidae) formen un grup de macrolepidòpters molt heterogeni. Amb nombroses excepcions, aquestes són les característiques més o menys generals: nocturnes, ales amples i vivament colorejades, òrgan timpànic al tòrax, espiritrompa atrofiada, postes nombroses i agrupades. El nom de la família al·ludeix a la pilositat de les eruges, que recorda l’aspecte de la pell dels óssos (arctos, ós). Les erugues típiques tenen llargs pèls negres, són hivernants, es traslladen amb gran rapidesa i fabriquen un capoll amb seda i pèls. És una família molt important (comprèn 10 000 espècies aproximadament), especialment a Amèrica del Sud, representada per 80 i 45 espècies a Europa i als Països Catalans, respectivament. N’indicarem les tres subfamílies presents al nostre país (que alguns autors eleven a categoria de família).
Els arctins (Arctiinae) són els que s’ajusten més a les característiques indicades, presenten colors aposemàtics, premonitoris de la seva toxicitat, i algunes femelles tenen les ales atrofiades o són àpteres. Dels litosins (Lithosiinae), hi ha autors que en separen determinats gèneres en una família a part, la dels endròsids (Endrosidae), però, en qualsevol cas, no és sinònim de la subfamilia. Són els més petits dels àrctids, amb ales críptiques, de color de palla (les anteriors, llargues i estretes, cobreixen tot el cos, i semblen un tros de tija seca), les erugues mengen molses i líquens; els cal·limorfins (Callimorphinae) són els més grossos, diürns i amb una representació mínima als Països Catalans (tres espècies). Alguns autors els consideren amb categoria de família, amb el nom d’hípsids (Hypsidae,) perquè no tenen l’anomenat timbal (òrgan que emet ultrasons) que tenen tots els altres àrctids i els sintòmids. Hi ha espècies molt abundants, com Eilema deplana, Spiris striata, Arctia caja, A. villica, Diacrisia sannio, Phragmatobia fuliginosa i Callimorpha quadripunctaria.
Els sintòmids
Els sintòmids (Syntomidae) constitueixen una família de lepidòpters diürns principalment sudamericana, que ha rebut també els noms de ctenúquids (Ctenuchidae) i amàtids (Amatidae), i que determinats autors consideren com una subfamilia dels àrctids. Hom en coneix en conjunt al voltant d’unes 3000 espècies, set de les quals són europees. A la zona tropical, presenten mimetisme amb himenòpters agressius. Als Països Catalans només tenim dues espècies, una de les quals (Syntomis phegea) és sorprenentment mimètica amb el zigènid tòxic Zygaena ephialtes, amb el qual és molt fàcil confondre’l.
Els noctúids: cucs grisos o dormidors, barrinadors i afins
En línies generals els noctúids (Noctuidae) constitueixen un grup més homogeni que el dels àrctids, però, atesa la gran quantitat d’espècies que reuneix (unes 40 000; és a dir, més que la majoria d’ordres d’insectes), és normal que hi hagi diversitat d’opinions quant a la col·locació de determinats gèneres i subfamílies. Seguidament, indiquem un seguit de caràcters aplicables a la majoria de tàxons.
Son macrolepidòpters d’activitat nocturna, d’on ve el nom de la família. Aspecte més aviat robust i pilós. Al tòrax tenen òrgans timpànics. L’abdomen és mes estilitzat i d’un sol color, apagat. Les antenes són filiformes, i en alguns mascles, pectinades. L’aparell bucal és ben desenvolupat, amb palps de vegades molt llargs. Les ales anteriors són llargues i estretes, amb dissenys críptics; les posteriors són més arrodonides i més clares (blanques, grises, marrons, etc.) de vegades de colors vius (vermell, taronja, groc, etc.) que exhibeixen en ésser atacats per tal de sorprendre l’intrús. De vol ràpid i rectilini, algunes espècies són grans migradores. La grandària és molt variable: des dels gèneres Nycteola, Eublemma i Porphyrinia que semblen microlepidòpters, al gènere Thysania, que inclou el lepidòpter de més envergadura alar (T. agripina, de 30 cm).
Els noctúids són la família més evolucionada dels lepidòpters, i tant l’eruga com l’adult i la crisàlide han assolit un elevat grau d’adaptació: el seu nombre d’espècies i la magnitud de les poblacions i, per tant, el seu grau d’incidència ecològica, els converteixen en la família més universal i important de tots els lepidòpters. En molts casos, com ara el d'Apopestes spectrum l'eruga passa desapercebuda gràcies al camuflament que li proporciona la coloració críptica i la disposició totalment aplanada sobre el substrat.
Les postes solen ser nombroses i agrupades. Les erugues, amb 3, 4 o 5 parells de pseudopodis, normalment són glabres. Per a crisalidar s’enterren, però sense fer embolcall pupal de seda. Moltes espècies són plagues forestals, però sobretot agrícoles, com el barrinador de la carxofa (Gortyna xanthenes), un dels barrinadors del blat de moro (Sesamia nonagrioides) i els cucs grisos de les hortes, nom popular que engloba nombroses espècies dels gèneres Spodoptera, Autographa i altres.

Un dels noctuïns, Noctua pronuba.
Jordi Vidal
Es tracta de la família més nombrosa i amb més importància ecològica dels lepidòpters, i la que té més representants als Països Catalans (700, de les 1200 conegudes a Europa). És cosmopolita i colonitza els més diversos biòtops. Donada la gran quantitat de subfamílies que hi ha, no les podem descriure. Tan sols assenyalarem els dos grans grups amb diferent nervació alar: els trífids i els quadrífids (aquests tenen un nervi més a les ales posteriors).
De la subfamília dels plusins destaquen Chrysodeixis chalcites, Bena prasinana i Autographa gamma, un dels lepidòpters catalans més estesos i abundants, que ocasiona greus plagues agrícoles. La seva eruga, així com també les d’altres gèneres, com Heliothis, Mamestra, etc., són conegudes amb el nom de cucs dormidors o cucs grisos. Altres espècies importants són Mythimna vitellina, Acronicta psi, Heliothis armigera i Catocala nymphagoga.
Els noctuïns són la subfamilia més nombrosa dels noctúids. Entre ells hi ha Noctua pronuba, de 57 mm d’envergadura alar; Peridroma saucia i el gènere Agrotis, que inclou diverses espècies de cucs grisos molt perjudicials per a l’agricultura com A. segetum, A. exclamationis i A. ipsilon, una de les plagues més polifagues i difícils de combatre.
Finalment, assenyalarem dues antigues famílies, actualment integrades com a subfamílies dels noctúids. Els nolins (Nolinae), antigament inclosos dins dels àrctids i considerats durant molt temps com una família independent, són petites papallones que volen de nit, de les quals hom en coneix una dotzena d’espècies als Països Catalans. Les eruges mengen líquens. Els dilobins (Dilobinae) constitueixen un grup monospecífic, peculiar per l’especial estructura de l’òrgan timpànic i la rara combinació de la nervació alar, la genitàlia i la resta de la morfologia. Per aquesta raó, l’espècie (Diloba caeruleocephala) s’ha anat incloent a les famílies dels notodòntids, tiatírids, bombícids i limàntrids. Al final, com sol passar en aquests casos, s’ha creat una família a part. Tanmateix, recentment, sembla demostrat que es tracta d’un noctúid força modificat. Alguns autors fins i tot situen aquesta espècie dins la subfamilia dels cucul·lins (Cuculliinae), al costat del gènere Episema, tot i que possiblement es tracta d’un panteiní (Pantheinae). L’eruga menja ametllers i altres arbres fruiters, i pot constituir plaga als Països Catalans. A l’inrevés, assenyalarem que una altra subfamilia, la dels herminins (Herminiinae) ha estat recentment separada dels noctúids i és considerada com una família independent per alguns autors.