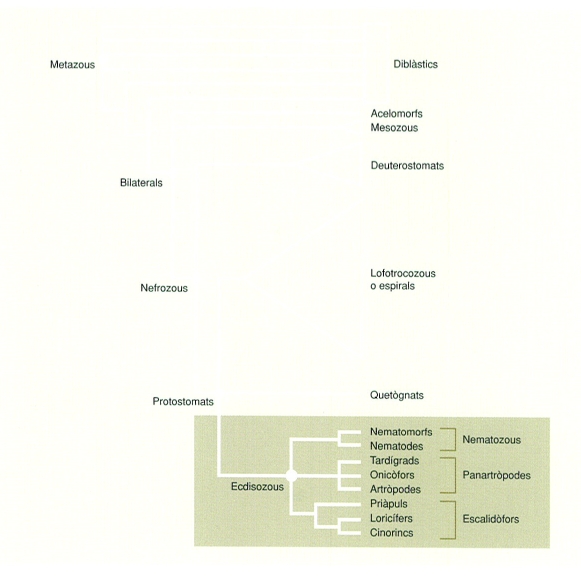
Esquema consensuat de les relacions externes dels artròpodes, amb especial atenció als fílums més estretament relacionats, els quals constitueixen, juntament amb els artròpodes, el gran clade protostomat dels ecdisozous. Els protòstoms inclouen també els lofotrocozous (anomenats per alguns autors espirals) i els quetognats, un fílum de posició encara incerta. Els deuteròstoms i els protòstoms constitueixen un llinatge evolutiu que hom anomena nefrozous, per la presència en aquests grups dòrgans excretors. Dos petits grups, els acelomorfs, antigament inclosos en els platihelmints, i els mesozous constitueixen, juntament amb els nefrozous, el llinatge dels bilaterals.
Original de l’autor, a partir de fonts diverses.
“La nova filogènia animal", així titulaven el desaparegut científic francès André Adoutte (1947 - 2002) i alguns dels seus col·laboradors un article publicat l’any 2000 en la prestigiosa revista Proceedings of the National Academy of Sciences. Amb aquest terme, els autors feien referència a les noves, i a vegades revolucionàries, afinitats evolutives dels principals llinatges animals sorgides de l’aplicació de les tècniques moleculars (en especial, la seqüenciació del DNA) a l’estudi de les relacions genealògiques entre els éssers vius. El tret de sortida de la incorporació del DNA a la inferència filogenètica s’havia donat anys abans, el 1988, amb la publicació d’un treball de Katharine Field i col·laboradors en la revista Science en què s’utilitzava la seqüència nucleotídica del gen de la subunitat petita ribosòmica (18S) per a establir, ni més ni menys, la filogènia del regne animal. Com és freqüent quan apareixen i s’adopten noves tecnologies, hi va haver un temps d’optimisme il·limitat al voltant de la capacitat de les dades moleculars per a resoldre l’arbre de la vida, i es va fer un ús indiscriminat i poc crític d’aquestes dades, depreciant en molts casos l’evidència proporcionada per altres fonts de caràcters, com ara la morfologia. Avui dia, ha passat prou temps perquè es comencin a conèixer les limitacions i els problemes metodològics inherents a l’ús del DNA com a eina filogenètica; alhora, s’ha acumulat l’experiència per a millorar els mètodes d’anàlisi i desenvolupar-ne de nous. Cal no oblidar que els fonaments de bona part del que se sap avui de l’arbre de la vida dels animals es van establir a partir dels estudis anatòmics morfològics i embriològics, tant d’organismes actuals com fòssils, duts a terme els darrers segles. És ben cert, però, que les dades moleculars han proporcionat una eina excel·lent per a afinar algunes d’aquestes relacions, per a resoldre aspectes controvertits i per a revelar noves relacions mai abans proposades, i que han obligat a reexaminar de forma crítica l’evidència morfològica que les sustenta. En aquest punt, convé desfer un equívoc freqüent quan hom parla dels avantatges de les tècniques moleculars. Sovint, es critiquen les dades morfològiques perquè suposadament sustenten hipòtesis filogenètiques que s’han demostrat errònies sota la llum de la filogènia molecular. Tanmateix, aquesta comparació és injusta, ja que el que es compara no és tant els tipus de dades com l’ús d’una metodologia quantitativa, que empra algoritmes informàtics, envers propostes més o menys subjectives, sorgides de l’intel·lecte d’un especialista en un grup d’organismes. L’anàlisi de dades morfològiques convenientment tractades per mètodes similars als emprats amb dades moleculars pot donar lloc a hipòtesis més fàcilment reconciliables amb la nova filogènia.
A grans trets, es poden distingir quatre fases al llarg dels gairebé vint anys d’ús de tècniques moleculars en filogènia, de límits difosos i no necessàriament successives en el temps. La primera fase correspondria a l’ús d’un sol gen, generalment el 18S ribosòmic; la segona, a l’anàlisi combinada de diferents gens, tant ribosòmics com proteics, bé nuclears o mitocondrials. Paral·lelament, es començaren a combinar dades moleculars amb altres de morfològiques mitjançant l’anàlisi simultània de matrius mixtes. Finalment, i gràcies a l’avenç tecnològic produït en la seqüenciació del DNA, es desenvolupà la filogenòmica, és a dir, l’anàlisi de genomes complets, o gairebé complets.
La posició dels artròpodes dins l’arbre de la vida
Els anèl·lids i els artròpodes han estat tradicionalment inclosos en un llinatge evolutiu comú, conegut com articulats (Articulata), basant-se en la segmentació del cos, present en ambdós grups. De fet, dins l’esquema evolutiu proposat per alguns autors que a la segona meitat del segle passat defensaven l’origen polifilètic (és a dir, independent) dels principals grups d’artròpodes, els anèl·lids compartien un avantpassat comú amb els grups que presenten apèndixs unirramis (d’un sol eix, per exemple, insectes, centpeus i milpeus). Al final dels anys vuitanta i el començament dels noranta, les primeres filogènies moleculars, basades en l’anàlisi del gen ribosòmic nuclear 18S, indicaven clarament que els anèl·lids no estaven estretament emparentats amb els artròpodes, sinó amb el mol·luscs, unes afinitats que van ser confirmades per anàlisis quantitives posteriors de caràcters morfològics.
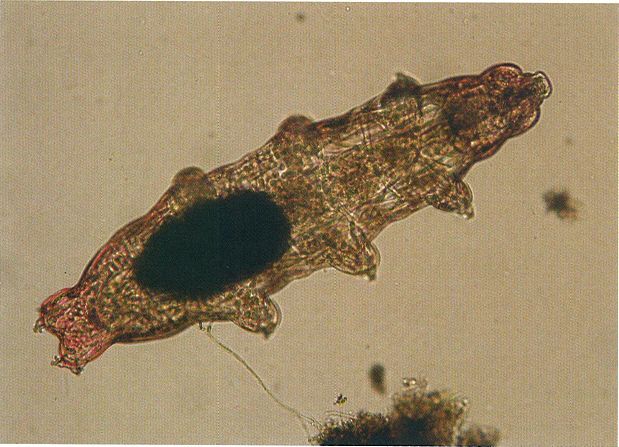
Exemplar de tardígrad recol·lectat al Maresme. Tot i que clàssicament els tardígrads han estat considerats un grup força relacionat amb els artròpodes i els onicòfirs, amb els quals constituirien el grup dels panartròpodes, algunes dades filogenòmiques apunten la possibilitat que els tardígrads puguin estar més directament emparentats amb els nematodes. Seran necessaris, per tant, més estudis per a determinar la seva posició definitiva.
Eduardo Mateos
L’any 1997, utilitzant el gen 18S sobre una representació més àmplia de fílums d’animals, es va poder demostrar l’existència d’un llinatge evolutiu que incloïa aquells en els quals es dóna l’ècdisi, és a dir, el creixement mitjançant mudes de la cutícula externa. El nou llinatge va ser anomenat ecdisozous (Ecdysozoa) i definit com el grup que inclouria l’avantpassat comú i tots els descendents d’artròpodes, tardígrads, onicòfors, nematodes i nematomorfs, priàpuls i cinorincs. Curiosament, l’existència dels ecdisozous ja havia estat avançada per estudis morfològics del final del segle XIX i inicis del XX i també obtinguda mitjançant l’anàlisi quantitativa de dades morfològiques. Aquest llinatge evolutiu ha estat posteriorment corroborat per nombrosos estudis moleculars i anàlisis combinades de dades moleculars í morfològiques. El complex de gens Hox, que estan involucrats en la diferenciació regional del cos dels animals, proporciona una font addicional de l’origen evolutiu comú dels ecdisozous, ja que certs gens d’aquest complex (Ubx, abd-A i Abd-B) mostren un pèptid específic en el grup i, alhora, a tots els manca el gen ParaHox Xlox. A mitjan dècada del 2000, però, els primers estudis filogenòmics (aquells que utilitzen la informació de genomes complets per a la inferència filogenètica) van suggerir que els artròpodes estarien més relacionats amb els vertebrats que no pas amb els nematodes (hipòtesi anomenada Coelomata, per la presència en ambdós grups de cavitats celomàtiques). Tanmateix, posteriorment s’ha demostrat que els resultats d’aquests estudis pioners eren un artefacte produït per l’efecte d’atracció de les rames llargues (anomenat així l’error que succeeix quan les branques d’un arbre filogenètic amb moltes substitucions tendeixen a ajuntar-se independentment de si els dos organismes estan realment relacionats o no) i l’esbiaix en la composició nucleotídica. Una de les principals dificultats per a l’acceptació universal dels ecdisozous ha estat la dificultat d’establir les característiques morfològiques que els defineixen envers altres llinatges. S’han proposat, juntament amb la muda i el seu control hormonal (la muda és regulada per l’acció d’hormones esteroides), l’estructura particular de la cutícula i la presència en alguna de les seves capes d’un polisacàrid característic. Cap d’aquests trets, però, és exclusiu dels artròpodes; les cutícules i les hormones ecdisteroides, per exemple, estan àmpliament distribuïdes al llarg dels metazous, i d’altra banda, el reemplaçament de la cutícula també és present en organismes no relacionats, com alguns poliquets o les sangoneres. Alguns autors argumenten que no és tant la presència d’aquests caràcters com el fet que tots ells s’expressen d’una manera particular característica dels ecdisozous. Altres caràcters proposats per definir aquest grup són l’absència de cilis en les cèl·lules epidèrmiques, la boca terminal i el cervell circumesofàgic. L’existència dels ecdisozous té una forta rellevància evolutiva, ja que suggereix que l’avantpassat dels bilaterals devia ser segmentat i, per tant, un organisme més complex del que generalment s’havia acceptat. D’altra banda, el fet que dos dels principals organismes models en biologia com són l’artròpode Drosophila melanogaster i el nematode Caenorhabditis elegans, estiguin més estretament emparentats del que es pensava, limita la universalitat dels estudis d’aquests organismes per explicar la forma i funció dels animals bilaterals.
Les dades filogenètiques més recents mostren que el que tradicionalment s’anomenaven asquelmints (cucs sense cavitats o només amb cavitats primàries o pseudoceloma) inclou en realitat dos grups evolutius independents, un relacionat amb els lofotrocozous (o espirals) i l’altre relacionat amb els artròpodes, amb els quals constitueixen els ecdisozous. Dins aquest últims trobem els fílums dels priàpuls, els cinorincs, els loricífers, els nematodes i els nematomorfs. Alguns autors proposen que aquests fílums constituirien un grup evolutiu amb un ancestre comú, que anomenen Cycloneuralia o Introverta, amb referència al fet que en tots s’observa un cervell circumesofàgic i una probòscide evaginable, respectivament. Els resultats de les anàlisis moleculars, i la combinació d’aquestes amb caràcters morfològics, són ambigus envers aquesta associació. En canvi, els grups dels escalidòfors (Scalidophora) o cefalorincs (Cephalorhyncha), que inclouria priàpuls, loricífers i cinorincs, i els nematozous (Nematozoa o Nematoida), els nematodes i els nematomorfs, reben més suport filogenètic. Els escalidòfors es caracteritzen perquè tenen la probòscide evaginable recoberta d’espines i papil·les, i dos anells de músculs retractors, mentre que en els nematomorfs la cutícula no és formada per quitina sinó per col·lagen, no tenen musculatura circular, els cordons nerviosos són ventrals i/o dorsals, tenen cloaca i els espermatozous han perdut el flagel.
L’estreta relació entre tardígrads, onicòfors i artròpodes, llinatge conegut amb la denominació de panartròpodes (Panarthropoda), és ben refermada per la presència d’apèndixs seriats ventrolaterals, amb musculatura extrínseca i intrínseca, i ungles terminals. La major part d’anàlisis filogenètiques morfològiques, i aquelles que inclouen dades paleontològiques i moleculars, corroboren l’origen comú d’aquests grups, tot i que les relacions entre ells són objecte de discussió. Mentre que alguns autors defensen que els tardígrads serien el grup germà dels artròpodes, amb els quals compartirien els apèndixs articulats amb musculatura intrínseca, un cervell de tres segments (caràcter aquest força discutit) i musculatura estriada, d’altres proposen que són els onicòfors els que compartirien un avantpassat exclusiu amb els artròpodes, tal com indiquen el sistema circulatori hemocèlic obert, amb un cor dorsal i ostíols, la musculatura metamèrica als apèndixs, les comissures en els nervis ventrals i els nefridis celomàtics. Els estudis filogenòmics més recents reforcen el parentiu d’onicòfors i artròpodes. Cal destacar que aquests mateixos estudis, i altres basats en pocs gens, revelen que els tardígrads podrien no estar emparentats amb els artròpodes o els onicòfors, sinó amb els nematozous. Tot i que aquesta relació dista molt de ser acceptada, és interessant destacar l’existència de caràcters similars en tardígrads i nematodes, com ara l’estructura de l’aparell bucal i la faringe, i d’alguns òrgans sensorials, i com també la presència d’un nombre de cèl·lules fix en l’adult (fenomen anomenat eutèlia). Tanmateix, hi ha autors que interpreten aquestes semblances bé com a caràcters ancestrals dels ecdisozous o bé com a convergències resultants del procés de miniaturització d’aquests grups.
La filogènia dels artròpodes
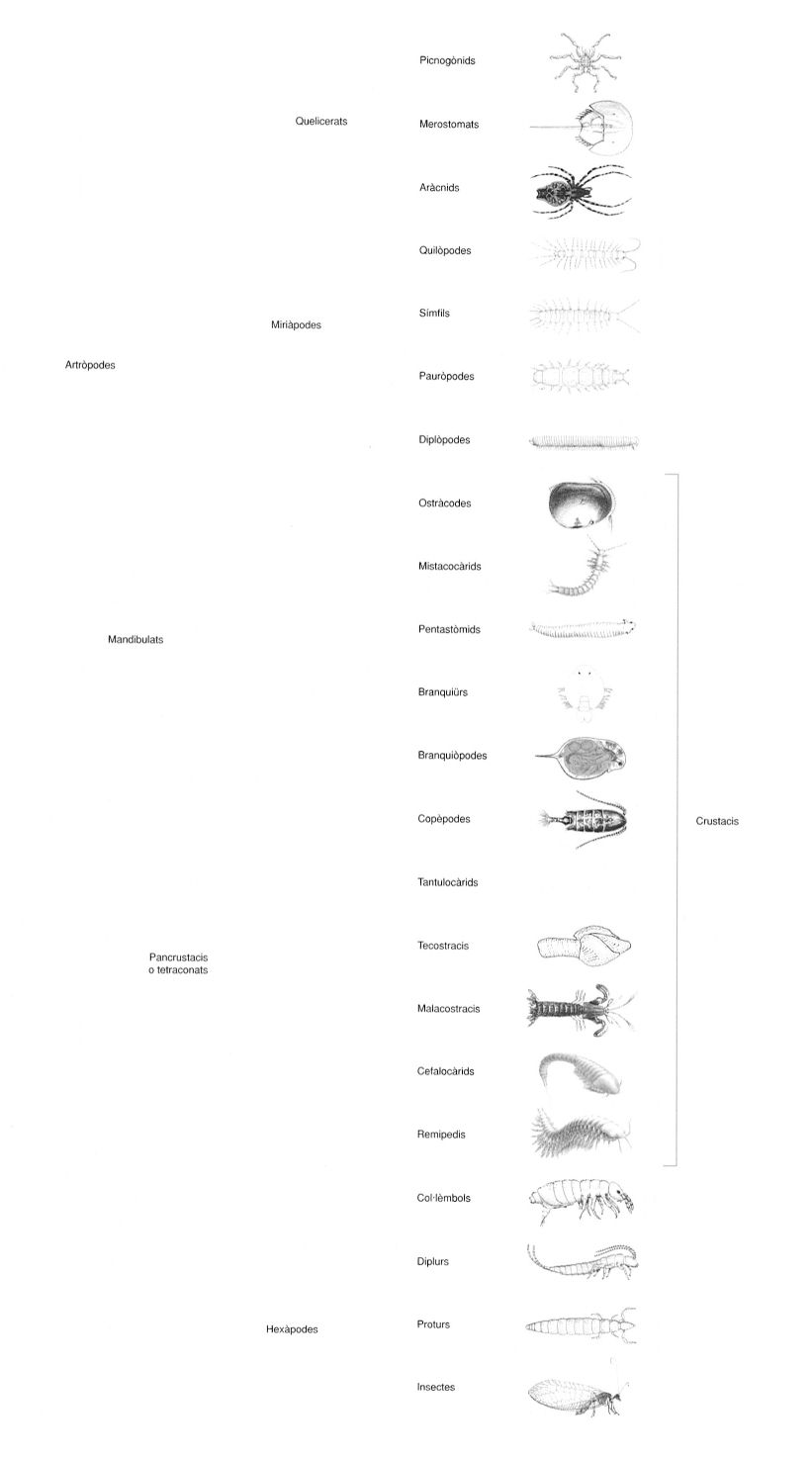
Relacions filogenètiques entre els principals llinatges d’artròpodes existents, d’acord amb el coneixement actual consensuat pels especialistes. Malgrat els interrogants que encara hi ha, les noves tecnologies combinades amb la ingent quantitat de dades acumulades al llarg de segles d’estudi, han permès fer un llarg camí. Tanmateix, la filogenètica és una ciència històrica i, per tant, subjecta a una constant revisió sota noves fonts d’evidència, a vegades conflictives a vegades ambigües, però que esperonen els científics a seguir treballant per assolir el somni de Darwin: reconstruir l’arbre de la vida a la Terra.
Original de l’autor, a partir de fonts diverses / Jordi Corbera, Amadeu Blasco, Marisa Bendala, Gustavo Hormiga, Miquel Alonso, Eduardo Saiz i Agnès Perelló.
Els artròpodes actuals han estat classificats tradicionalment en quatre grans grups, generalment amb categoria taxonòmica de subfílums, basant-se en les principals regions corporals (tagmes) i els apèndix característics associats; quelicerats, miriàpodes, crustacis i hexàpodes. Un cinquè grup, els dels trilobits, s’hauria extingit al final del Permià, després d’haver estat els artròpodes dominants al llarg del Paleozoic. Les relacions filogenètiques entre aquests quatre grans grups han estat font de controvèrsia. D’una banda, hi ha qui defensa l’existència de dues línies evolutives independents: la dels unirramis (Uniramia), que inclouria els miriàpodes i els hexàpodes, i, segons d’altres, els onicòfors, que comparteixen el fet de tenir un únic parell d’antenes (els crustacis en tenen dues i els quelicerats cap) i els apèndixs formats per un sol eix, i els TCC (per les sigles en anglès de trilobits, quelicerats i crustacis), també anomenat dels esquizoramis (Schizoramia), pels apèndixs de dos eixos (birramis). Aquesta hipòtesi filogenètica és una de les teories polifilètiques anteriorment esmentades, les quals defensaven un procés convergent d’artropodització a partir de llinatges evolutius no directament emparentats. Tanmateix, aquesta visió no va ser mai basada en anàlisis filogenètiques quantitives, sinó en argumentacions sobre grups de caràcters particulars. El gros de l’evidència filogenètica actual, morfològica i molecular, referma de manera inequívoca l’origen comú i únic dels artròpodes. En l’actualitat, els principals punts de conflicte pel que fa a les relacions entre els principals llinatges d’artròpodes són tres: la posició dels picnogònids, la posició dels miriàpodes i la relació entre crustacis i hexàpodes.
Els picnogònids han estat considerats tradicionalment quelicerats perquè disposen d’un parell d’apèndixs anteriors en forma de pinça, els quelí-fors, que han estat interpretats com homòlegs dels quelícers de la resta de quelicerats. Tanmateix, algunes anàlisis filogenètiques de dades morfològiques i moleculars suggereixen que els picnogònids podrien ser el grup germà de la resta d’artròpodes actuals. Aquesta hipòtesi va rebre la corroboració d’un estudi neuroanatòmic embriològic, que indicava que els quelífors probablement són innervats pel protocervell (preorals), a diferència de quelícers i antenes, que ho són pel deutocervell, i, per tant, permetia posar en dubte que es tractés d’apèndixs homòlegs. Altrament, l’existència d’apèndixs preorals ha portat alguns autors a suggerir que els picnogònids podrien ser els únics representants actuals d’un grup extingit d’artròpodes càmbrics, caracteritzat per la presència d’un parell d’apèndixs frontals molt desenvolupats i coneguts per això com el llinatge dels grans apèndixs. Ara bé, aquesta evidència ha estat posada en dubte per estudis posteriors, que han demostrat, mitjançant l’expressió dels gens Hox i estudis neuroanatòmics d’una mostra més àmplia de picnogònids, l’homologia de quelífors i quelícers. A més, les anàlisis filogenètiques morfològiques que inclouen fòssils reinterpreten els grans apèndixs com a possibles quelícers. Els estudis filogenòmics recents corroboren la visió tradicional dels picnogònids com a quelicerats, i els situen com a grup germà de la resta de quelicerats (Euchelicerata).
Tots els miriàpodes, crustacis i hexàpodes presenten un parell d’apèndixs alimentaris propis anomenats mandíbules, un caràcter que s’ha fet servir per agrupar-los sota el nom de mandibulats. Tanmateix, un elevat nombre d’anàlisis moleculars han posat en dubte la validesa evolutiva d’aquest caràcter, i en canvi han donat suport a la idea d’una estreta relació entre miriàpodes i quelicerats, que formarien el grup dels anomenats paradoxòpodes (Paradoxopoda) o mirioquelats (Myriochelata).

Picnogònid procedent de l’oceà Antàrtic (illes Shetland del Sud), d’una espècie en què manquen els quelífors i els palps. A la imatge s’aprecia la trompa característica del grup i els petits apèndix anteriors oovígers, que s’utilizen per a tranportar la posta. Malgrat que alguns estudis moleculars han suggerit que serien el grup germà de la resta d’artròpodes actuals, les dades més recents confirmen la seva posició dins el llinatge dels quelicerats i corroboren, per tant, l’homologia de quelifors i quelícers.
Manuel Ballesteros
Aquesta relació, malgrat la sorpresa inicial que va suposar, atesa l’absència de caràcters morfològics que la reforcin, fou corroborada per estudis posteriors amb gens Hox i sobre la neurogènesi d’ambdós llinatges. Altres estudis, però, han demostrat que aquesta relació és probablement un artefacte metodològic, ja que segons quin grup extern de referència es fa servir s’obté el grup dels mandibulats o el dels paradoxòpodes. D’altra banda, les dades filogenòmiques donen un suport sòlid a la hipòtesi dels mandibulats, més coherent amb la morfologia.
Una de les relacions menys qüestionades a les classificacions tradicionals ha estat la de miriàpodes i hexàpodes. Ambdós llinatges formarien el grup dels atelocerats (Atelocerata) o traqueats (Tracheata), noms que fan referència a caràcters tals com la possessió d’un sol parell d’antenes, amb un segment intercalar sense apèndixs, o la presència de dendrotràquees, respectivament, a més de compartir tots dos grups els tubs de Malpighi com a òrgan d’excreció. Alguns esquemes filogenètics fins i tot anaven més enllà i proposaven que alguns grups de miriàpodes serien evolutivament més propers als hexàpodes que a altres miriàpodes, convertint els anteriors en un grup parafilètic. La irrupció de les filogènies moleculars va trasbalsar aquesta visió plantejant que eren el crustacis, i no el miriàpodes, el grup d’artròpodes més íntimament relacionats amb els hexàpodes. Aquesta nova agrupació, originalment batejada amb el nom de pancrustacis (Pancrustacea), actualment és coneguda com a tetraconats (Tetraconata), en referència a l’estructura comuna dels ommatidis d’ambdós grups, que inclou un con cristal·lí dividit en quatre parts. El gros de l’evidència molecular actual, incloent-hi dades filogenòmiques, referma els tetraconats sense cap mena de dubte. D’altra banda, els fòssils tenen un paper important, ja que incloure’ls en les anàlisis morfològiques reforça els traqueats (Tracheata) i excloure’ls dóna suport als tetraconats.

Els remipedis, juntament amb un altre petit llinatge de crustacis marins (els cefalocàrides), són vistos actualment com a possibles grups germans dels hexàpodes, és a dir, més propers als hexàpodes que no a la resta de llinatges de crustacis. Malgrat les dades que s’han anat acumulant a partir de les anàlisis morfològiques i moleculars, encara no hi ha consens sobre les relacions entre els principals llinatges de crustacis.
Texas A&M University / Thomas lliffe
Els crustacis són l’únic gran llinatge d’artròpodes, l’origen de les relacions internes dels quals no està consensuat encara avui dia. Les anàlisis moleculars de conjunts de gens coincideixen amb les anàlisis morfològiques quantitatives a rebutjar bona part dels grups taxonòmics tradicionals. Així, l’evidència actual suggereix que els antics maxillòpodes són un grup artificial, que els peus de cabra i glans de mar estan força més relacionats amb els malacostracis (galeres, escamarlans o porquets de Sant Antoni) i que ostracodes, mistacocàrides i branquiürs formarien un llinatge comú independent. D’altra banda, la possibilitat que els crustacis no siguin un llinatge evolutiu exclusiu, sinó que alguns grups siguin evolutivament més propers als hexàpodes que no pas altres, és un resultat recurrent de les anàlisis moleculars, tot i que hi ha estudis que es contraposen atès que donen suport a diferents candidats a grup germà dels hexàpodes. En aquest sentit, la possibilitat que el branquiòpodes siguin el grup germà dels hexàpodes, ha dut alguns autors a suggerir un escenari evolutiu en què els hexàpodes serien crustacis que van colonitzar el medi terrestre mitjançant un estadi intermedi adaptat a aigües continentals, representat pels actuals branquiòpodes. Malauradament per a aquesta hipòtesi, les dades filogenòmiques més recents situen els cefalocàrids i els remipedis (Xenocarida), dos petits llinatges de crustacis marins, com el grup germà dels hexàpodes (i no els branquiòpodes). D’altra banda, cal destacar que totes les dades moleculars apunten que els pentastòmids, que pel seu aspecte aberrant han estat considerats com un fílum independent més o menys relacionat amb artròpodes, estan fortament emparentats amb els branquiürs, un grup de crustacis ectoparàsits de peixos. Tanmateix, aquestes dades estan en conflicte amb el registre fòssil, que indica una edat molt antiga dels pentastòmids, que ja serien presents al Cambrià superior, la qual cosa sembla més compatible amb la seva separació primerenca de la resta d’artròpodes.

Exemplar de col·lèmbol de l’espècie Bilobella aurantica, provinent de Collserola (Barcelonès). La posició dels col·lèmbols dins l’arbre filogenètic dels artròpodes ha estat molt controvertida, fins al punt que s’ha arribat a dubtar que s’haguessin de considerar dins el llinatge dels hexàpodes, si bé actualment es tendeix a donar validesa a l’origen comú de tots els hexàpodes, incloent-hi els col·lèmbols, i a la inclusió d’aquests últims juntament amb els proturs i els diplurs en el llinatge dels entògnats.
Eduardo Mateos
Els hexàpodes han estat tradicionalment separats en dos grups segons el tipus de la seva càpsula cefàlica. Els entògnats (Entognatha: proturs, diplurs i col·lèmbols) tenen les peces bucals cobertes per la càpsula cefàlica, mentre que en els ectògnats (Ectognatha: peixets de coure i de plata, i els insectes amb ales) les peces bucals resten descobertes. L’origen evolutiu comú dels ectògnats, els insectes en un sentit ampli, no ha estat posat en dubte, però les relacions del grups d’entògnats són controvertides. Les anàlisis morfològiques i moleculars, independents o combinades, reforcen que proturs i diplurs constitueixen un llinatge evolutiu propi, que s’anomena nonoculats (Nonoculata). Ara bé, l’anàlisi de genomes mitocondrials complets qüestiona que els col·lèmbols tinguin un origen comú tant amb els nonoculats com amb la resta d’hexàpodes. Aquestes dades suggereixen que els col·lèmbols haurien evolucionat a partir d’un llinatge de crustacis independent. Aquesta hipòtesi té importants implicacions evolutives, ja que suposa que l’adquisició del pla corporal dels hexàpodes hauria tingut lloc en dues fases, probablement com a resultat de l’adaptació independent al medi terrestre. Aquests resultats, però, han estat fortament criticats, especialment per la metodologia emprada i pel limitat contingut informatiu dels caràcters analitzats. Anàlisis recents que utilitzen més de 60 gens proteics nuclears reforcen, però, l’origen únic dels hexàpodes i donen també suport al llinatge dels entògnats.