El gènere cosmopolita Artemia ha estat representat a la conca mediterrània només per poblacions de l’espècie A. salina i poblacions de soques partenogenètiques diploides i tetraploides, fins que va arribar l’artèmia americana (A. franciscana), que ha esdevingut invasora i amenaça d’eliminar per competència les espècies i les soques autòctones.
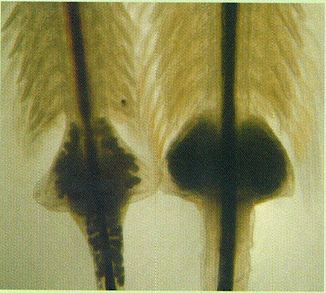
Detall de l’ovisac de femelles d’Artemia salina (a l’esquerra) i A. franciscana (a la dreta). En la primera s’aprecia la cintura que separa l’ovisac, més aviat triangular, del tòrax, i en la segona, la forma de l’ovisac més arrodonida i amb dos vèrtexs acabats en punta. S’hi veu també la diferent robustesa de l’abdomen de les dues femelles.
Francisco Amat.
Encara que els individus de les diferents espècies d’Artemia són molt semblants morfològicament en les poblacions silvestres (probablement per influència de l’alta salinitat del medi), els exemplars cultivats en condicions estandarditzades de laboratori permeten establir diferències que, un cop identificades, es poden arribar a reconèixer en els silvestres. Les diferències entre l’espècie bisexual autòctona A. salina i la invasora A. franciscana tenen a veure amb la forma general del cos i amb la forma de l’ovisac de la femella adulta. A més, els mascles d’A. franciscana presenten una espina a la part interna de la base del penis, que manca en els d’A. salina.
Les espècies del gènere Artemia són totes molt halotolerants, i aquesta adaptació els ha permès colonitzar hàbitats on tenen pocs predadors. Viuen en aigües de salinitat situada entre la de l’aigua de mar i la de la salmorra (300 g de sal/litre) i de composició química clorurada, sulfatada o carbonatada i d’aquí que habitin ecosistemes hipersalins litorals i continentals.
El paràmetre ambiental que més condiciona la distribució de les espècies d’Artemia és la temperatura. Les poblacions mediterrànies de l’espècie bisexual A. salina i de les soques partenogenétiques habituals poden conviure en un mateix lloc, però acostumen a mostrar una tendència estacional determinada pel règim tèrmic: les poblacions d’A. salina prefereixen les temperatures baixes de final d’hivern i inici de la primavera, mentre que les soques partenogenètiques es comporten a l’inrevés, de manera que s’estimen més l’estiu i la tardor, quan les poblacions d’A. salina ja han desaparegut. Tanmateix, A. franciscana es mostra més euriterma que les autòctones esmentades. El fet que hagi colonitzat tot el continent americà, de nord a sud, demostra una àmplia tolerància tèrmica, pròpia d’una gran plasticitat fenotípica, i aquesta tolerància fa que en un règim tèrmic com el mediterrani no trobi cap restricció estacional per a desenvolupar-se.

Aspecte dorsal de les femelles d’Artemia salina i A. franciscana. S’hi aprecien les tres parts anatòmiques pròpies dels anostracis: el cap, amb dues antènules i dos ulls pedunculats; el tòrax, amb onze parells de filopodis; i, al final, l’abdomen, compost de vuit segments –on destaca, just en els dos primers, l’ovisac on es desenvolupen els embrions-, i la furca terminal. S’hi pot notar la diferent robustesa de les femelles d’ambdues espècies, sobretot en el gruix de l’abdomen.
Francisco Amat.
L’estudi de l’eficàcia biològica o fitness comparada d’Artemia salina i A. franciscana mostra que aquesta darrera és més eficaç biològicament que no l’espècie autòctona amb la qual competeix. Tant en condicions experimentals com en el medi natural, l’espècie americana ha demostrat la seva precocitat a l’hora de madurar reproductivament i la capacitat de donar més descendència i amb més freqüència, durant un període reproductiu més llarg i amb un equilibri acurat entre la reproducció vivípara i ovípara, sempre amb fecunditat més elevada. Aquesta eficàcia biològica es tradueix en una capacitat reproductiva que supera de molt la de les altres espècies. És una de les característiques que determinen la condició d’invasores de les espècies que les posseeixen.
La biogeografia i la biodiversitat del gènere Artemia als Països Catalans ¡ a la resta de la península Ibèrica es coneix des dels anys vuitanta del segle passat. Se sap que les poblacions d’Artemia partenogenètica diploide de les salines de Gerri de la Sal (Pallars Sobirà) van ser malmeses per una inoculació d’A franciscana abans de desaparèixer. A les salines de la Trinitat, al delta de l’Ebre, es va trobar una de les poques poblacions d’Artemia partenogenètica tetraploide vivint en aquest tipus d’hàbitat (habitualment prefereix salines continentals), i avui aquesta salina és l’única del litoral llevantí envaïda per A. franciscana, cosa que ha provocat la desaparició de la soca partenogenètica esmentada. A les Balears (a les salines de Mallorca, Eivissa i Formentera) s’han descrit poblacions d’A. salina i de la seva soca partenogenètica diploide, les mateixes que es troben al País Valencià (a les salines alacantines de Santa Pola i Torrevella). També es poden esmentar aquí les salines de Calp, malauradament desaparegudes, on vivia una població partenogenètica diploide molt interessant.
En resum, actualment al litoral llevantí només es poden citar les salines del delta de l’Ebre com envaïdes per l’espècie Artemia franciscana. Cal anar-se’n fins a les salines de Màlaga, Cadis, Huelva i Portugal, en l’àmbit de la península Ibèrica, per a trobar poblacions ben establertes de l’espècie americana, per no parlar de les salines envaïdes de França i Itàlia.
La via d’arribada més probable d’Artemia franciscana és a través de cists que, produïts a la badia de San Francisco (Califòrnia) o al Gran Llac Salat de Utah (Estats Units), són transportats pels distribuïdors d’aqüicultura, sigui a petita escala (aquaris) o a gran escala (vivers d’alevins i larves de peixos, crustacis), si bé hi ha també la possibilitat que hagi estat introduïda expressament a les salines per aprofitar la capacitat que té de netejar les salmorres de partícules de matèria orgànica i així millorar la qualitat de la sal.
Un cop establertes les poblacions invasores, la seva dispersió és gairebé inevitable: els ocells aquàtics que freqüenten els medis hipersalins en transporten els quists a les potes, les plomes o dins l’aparell digestiu, i els dipositen en defecar en un aiguamoll salí prop o lluny del lloc de partida.
L’efecte més important de la introducció d’Artemia franciscana és l’aniquilació de les poblacions d’espècies autòctones. Des d’un punt de vista ecològic, s’hi poden associar conseqüències importants sobre espècies de paràsits d’ocells aquàtics (cestodes, per exemple), per als quals l’artèmia actua d’hoste intermediari. Encara no es pot avaluar l’impacte de la substitució d’una espècie per l’altra en l’alimentació dels ocells aquàtics. Se sap, però, que les artèmies parasitades de poblacions autòctones es fan més visibles i per tant són més vulnerables a la predació, cosa menys accentuada en l’artèmia americana. Així, la substitució d’una espècie per l’altra pot significar per a la majoria dels ocells que freqüenten els ecosistemes aquàtics hipersalins una pèrdua del seu aliment bàsic.
A continuació es presenta la taula de vida que compara l’eficàcia biològica (amb dades de l'autor), o fitness, de les espècies autòctones i les introduïdes a partir de paràmetres que representen la capacitat de competir en condicions ambientals determinades. Per a Artemia franciscana es van utilitzar parelles (♂ + ♀), i per a la partenogenètica, només femelles. Quan els paràmetres són més favorables, l’espècie està més capacitada per a competir amb les altres i perpetuar les seves poblacions en detriment de les altres, i es diu que aquesta espècie és més eficaç biològicament. La taula mostra que és el cas d’A. franciscana. * Condicions de cultiu: 24 oC de temperatura, 80-90 gl-1 de salinitat, medi nutritiu: cultiu microalgal de Tetraselmis i Dunaliella (1:1), fotoperíode: 12:12.
| TAULA DE VIDA | Artemia partenogenètica (tetraploide) N = 16 | Artemia franciscana N = 20 |
|---|---|---|
| Període prereproductiu (dies entre el naixement i la primera posta de la femella) | 23 ± 1,20 | 17,5 ± 0,9 |
| Període reproductiu (dies entre la primera i l’última posta abans de morir) | 39 ± 21 | 46 ± 30 |
| Nombre total de postes (totes les postes produïdes durant el període reproductiu) | 7 + 3,3 | 11 ± 6,2 |
| Dies entre postes (dies entre dues postes successives) | 6,4 ± 0,6 | 4,6 ± 0,4 |
| Descendència total (nauplis vivípars –vius. morts i embrions abortius– i cists) | 1.060 ± 635 | 2.707 ± 2.162 |
| Descendència per posta (descendència total / nombre de postes) | 140 ± 28 | 221 ± 60 |
| Descendència per dia (descendència total / període reproductiu) | 26,3 ± 4,4 | 54,5 ± 11 |
| Percentatge de viviparisme (% de nauplis vivípars: vius. morts i embrions vivípars abortius en la descendència total) | 1,77 ± 7 | 43,2 ± 26 |
| Percentatge d’oviparisme (ídem per als cists) | 98,2 ± 7 | 56,8 ± 26 |
| Durada total de vida (dies del període prereproductiu + el reproductiu + el postreproductiu) (dies entre l’última posta i la mort) | 63 ± 21 | 73 ± 30 |