Metodologia
Durant les dècades de 1960 i 1970, per a la pesca de zooplàncton s’utilitzaven xarxes amb una malla de 200-250 µm, la mateixa mostra que es feia servir per a l’estudi dels diferents grups que formaven part del zooplàncton: és a dir, no solament els copèpodes sinó també els sifonòfors, els cladòcers, els amfípodes, les larves de decàpodes, etc., mentre que per a l’ictioplàncton es feia servir la xarxa semipelàgica IKMT (Isaac Kit Mid Water Trawl), amb una malla de 500 µm, que permetia una velocitat d’arrossegament d’uns sis nusos. Estudis posteriors sobre la selectivitat de la malla en la pesca de zooplàncton han demostrat que una xarxa de 100 µm captura un gran nombre de nauplis, copepodits i alguns copèpodes, mentre que, a l’altre extrem, una xarxa de malla de 250 µm pesca molt pocs nauplis i molts més copèpodes adults. Per tant, actualment l’estudi dels microcopèpodes (de talla compresa entre 0,20 i 0,60 mm) es fa amb xarxes de 70 a 100 µm, arrossegades a velocitats pròximes a 0,5 m/s, i això ha permès comprovar la importància quantitativa dels microcopèpodes de grups com, per exemple, els onceids (Oncaea, Monotula, Tricoma i Spinoncaea) i descobrir nombroses espècies noves.
D’altra banda, els darrers anys s’han introduït aparells de pesca molt sofisticats, entre d’altres, la MOCNESS (Multiple Opening/Closing Net and Environmental Sensing Systems), que requereixen l’ús d’un complex equipament instal·lat a bord (com ara fils elèctrics, comandaments dirigits per ordinador, etc.), que permeten conèixer les característiques físiques del medi (salinitat, temperatura, etc.) de les aigües estudiades, i també l’obertura i el tancament de les boques de la xarxa.
Morfologia
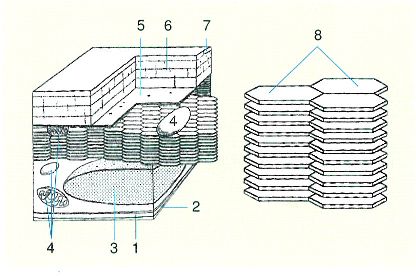
Formació laminar del tegument del prosoma dels mascles del gènere Sapphirina, a partir de l’estudi de la cutícula al microscopi electrònic. Els cristalls de guanina, més abundants en els mascles que en les femelles, són el material refringent dels copèpodes safrínids. D’acord amb el nombre de plaques de cada espècie, la llum que reflecteixen té longitud d’ona diferent, i, per tant, també colors diferents. 1 làmina basal, 2 membrana cel·lular, 3 nucli, 4 mitocondri, 5 membrana cel·lular, 6 procutícula, 7 epicutícula, 8 placa hexagonal.
Original de l’autor, a partir de Chae i Nishida, 1994.
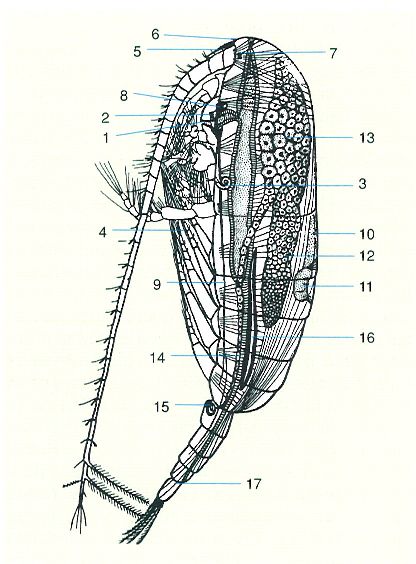
Anatomia interna dels copèpodes: femella de Calanus. 1 boca, 2 esòfag, 3 glàndula maxil·lar, 4 intestí, 5 filament rostral, 6 òrgan frontal, 7 ull mitjà, 8 cervell, 9 cordó nerviós ventral, 10 aorta, 11 cor, 12 ovari, 13 diverticle de l’ovari, 14 oviducte, 15 espermateca, 16 vesícula de l’oli, 17 anus.
Marshall i Orr, 1955.
Deixant de bana la morfologia general externa i interna dels copèpodes, convé afegir alguns detalls sobre els òrgans quimioceptors i mecanoceptors que complementen la funció ocular en captar diferents estímuls externs: l’òrgan de Gicklhorn, l’òrgan de Bellonci, l’òrgan pleural cefàlic, la gepa dorsal, les setes i les estetes.
Un altre aspecte d’interès és el fenomen de la bioluminescència o producció de llum a partir de reaccions químiques, que es manifesta a través dels porus del tegument. La distribució d’aquests porus pel cos de l’animal és idèntica en el mascle i la femella d’una mateixa espècie, però varia en cadascuna d’elles.
Les espècies que viuen a la zona fòtica es creu que poden distingir partícules amb més o menys resolució i diferents longituds d’ona (colors). Pel que fa a la distinció dels colors, en els safrínids es presenta iridescència i el material refringent són els cristalls de guanina. El contingut de guanina en el tegument del prosoma de certes espècies és molt més elevat en mascles que no en femelles. Al microscopi electrònic es pot veure que el tegument té una estructura laminar formada per hexàgons semblants a una bresca d’abella, amb diàmetre de cel·la d’un micròmetre (µm). El fet que aquestes plaques reflecteixin la llum s’explica per la teoria de la interferència per capes primes. Com que el nombre de sostres de plaques és diferent a cada espècie, la llum reflectida té diferents longituds d’ona (colors), que oscil·len entre 430 i 666 µm. Com que els safrínids viuen a la zona fòtica (generalment entre 50 i 150 m), cal pensar que la femella pot distingir pel color el seu congènere en el moment de la fecundació.
Pel que fa als copèpodes que viuen a la zona afòtica (grans profunditats, de 500 a 1.000 m), la unió dels dos sexes segurament té lloc pels diferents aspectes comentats: la distribució dels punts lluminosos, els òrgans quimiosensorials i mecanosensorials i les hormones sexuals.
La hidrografia i les poblacions de copèpodes
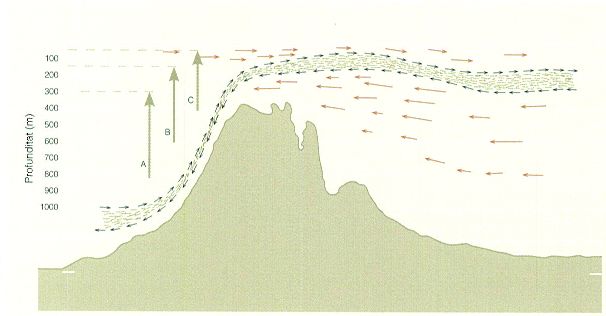
Intercanvi de poblacions de copèpodes a través de l’estret de Gibraltar. L’entrada d’espècies atlàntiques a la Mediterrània per l’estret és condicionada per l’amplitud de la migració nictemeral (diària). A l’esquema s’indica l’amplitud de la migració de tres grups de copèpodes: els del grup A es mouen entre 800 i 300 m; els del grup B ho fan des de 600 fins a 150; i els del grup C, des de 400 fins a 50-100 m. Aquest darrer grup és el més representat a les aigües de la Mediterrània.
Barreras & Creixell, a partir de Vives, Santamaria i Trepat, 1975.
És sabut que a la mar Mediterrània l’aportació d’aigua dolça pels rius no és suficient per a suplir la pèrdua d’aigua per evaporació, i això comporta una entrada d’aigües atlàntiques a través del canal de Gibraltar. Aquest fet determina que la circulació de la Mediterrània sigui del tipus estuàríc negativa: a l’estret, entre els 150 m de profunditat i la superfície, entra un milió de metres cúbics per segon (és a dir, un sverdrup) d’aigua atlàntica, mentre que en fondària, dels 150 m als 400 m de profunditat, en surt aigua mediterrània cap a l’Atlàntic en una quantitat menor (0,7 sverdrups/segon).
Les espècies que han donat lloc a la població actual de la mar Catalanobalear són en gran part d’origen atlàntic. Altres provenen de les aigües orientals, ja que aquest fenomen també es dóna entre la mar llevantina i la mar Roja, que és la via d’entrada de les espècies de la mar d’Aràbia a la Mediterrània. L’arribada d’espècies des de l’Atlàntic a la mar d’Alborán és condicionada per la migració vertical dels copèpodes a l’oest de Gibraltar, ja que només les espècies que en la seva migració pugen dels 400 m fins als 50-100 m són empeses pel corrent atlàntic i poden creuar l’estret de Gibraltar i arribar a la mar d’Alborán. Aquesta mar és el primer filtre de les poblacions que hi arriben, a causa del progressiu augment de la salinitat a què es veuen sotmeses. Moltes espècies atlàntiques que han entrat per Gibraltar es queden a la part meridional de la mar Mediterrània i no arriben a les aigües catalanes, i això fa que de manera general les espècies que es troben al sud de la Mediterrània s’hagin considerat indicadores d’aigües atlàntiques. Les que formen la població de la mar Catalanobalear, que en conjunt són unes 300 espècies, són les que han resistit l’augment de la salinitat.
Moviments i migracions
Malgrat que es consideren una part del plàncton, els copèpodes mostren patrons de moviment propis, fora del transport pels corrents, com ara les migracions verticals. Durant poques hores i sense que la massa d’aigua es mogui, els copèpodes són capaços de pujar centenars de metres d’una manera activa, i després es deixen caure (passivament, ja que són més densos que l’aigua de mar) a les profunditats d’on venien.
A part d’aquests moviments, n’hi ha d’altres, com són els de nutrició, que es fan per mitjà dels apèndixs bucals. L’animal es pot desplaçar de manera contínua o bé fent salts, segons si les antenes es mouen alternativament o bé juntes, com els rems d’una barca. Aquests moviments, i els de les potes (que es mouen de davant cap endarrere), fan que l’animal es desplaci amb petits impulsos.
Experiments d’alimentació han permès explicar la migració vertical o nictemeral dels copèpodes al llarg del dia. En els experiments, s’ha vist que individus adults d’una mateixa espècie posats en diferents recipients amb aigua de mar amb idèntica concentració d’aliment però sotmesos a il·luminacions de diferent intensitat, a una determinada intensitat de llum s’alimenten molt bé i, en canvi, en intensitats més altes o més baixes l’alimentació es redueix notablement. S’ha comprovat que aquestes espècies es troben a una profunditat on es registra la il·luminació més favorable, és a dir, el punt òptim de llum, i que quan, en disminuir la llum solar, el punt òptim va pujant dins la columna d’aigua, els copèpodes segueixen el mateix ascens, perseguint el punt òptim de llum. Quan arriba la foscor, els animals es troben en quantitats molt més grans als primers metres d’aigua que durant el dia, però al cap d’un cert temps tornen a baixar cap al fons. Dit d’una altra manera: la manca de llum, a efectes de l’alimentació, fa migrar cap a la superfície moltes espècies de copèpodes.
Aquests arguments no serveixen del tot quan es tracta d’esbrinar el comportament dels copèpodes que viuen a la zona afòtica. S’ha suposat que la migració també és deguda en part a l’alimentació, que dóna lloc a desplaçaments successius. Sembla que altres factors secundaris (a part dels bàsics de llum i alimentació), com ara la temperatura, el pH, la densitat de l’aigua, la salinitat, el contingut d’oxigen, el geotropisme, etc., tenen també una influència en la migració vertical. No cal dir que el comportament migratori és diferent segons les espècies, i també que varia dins la mateixa espècie segons el sexe.
Les migracions ontogèniques són les que es produeixen de manera més o menys estacional: durant certa part de l’any es troben prop de la costa i en altres èpoques s’allunyen cap a alta mar. És el cas, a la mar Catalana, de Calanus helgolandicus: quan es trenca la termoclina al final de la tardor apareixen abundantment individus adults a la zona nerítica prop de la costa, on mesos abans no n’hi havia cap exemplar. S’hi troben femelles adultes, que entre els mesos de gener i abril es reprodueixen tres vegades successives: al gener apareixen els primers nauplis, que ja són adults al final de febrer; la segona generació, al març-abril, i la tercera, a l’abril-maig, fins al juny. L’espècie, que és completament herbívora, coincideix amb les grans proliferacions de fitoplàncton, que s’inicien al final de gener o mitjan febrer. Al començament de l’estiu va desapareixent gradualment de la zona nerítica i migra cap a alta mar, de manera que a mesura que transcorre l’estiu van augmentant el nombre de copepodits entre els 500 i els 1.200 m. Cap al final de l’estiu i l’inici de la tardor, aquests copepodits mostren unes grans reserves lipídiques que els mantenen gairebé immòbils a nivells profunds en un estat d’hivernació i que són utilitzades per a la maduració dels òvuls. Així, a l’inici de l’hivern, quan la termoclina es trenca, els adults es troben prop de la costa gràcies a la invasió de les masses d’aigua d’alta mar, i el cicle torna a començar.
La classificació dels ordres de copèpodes
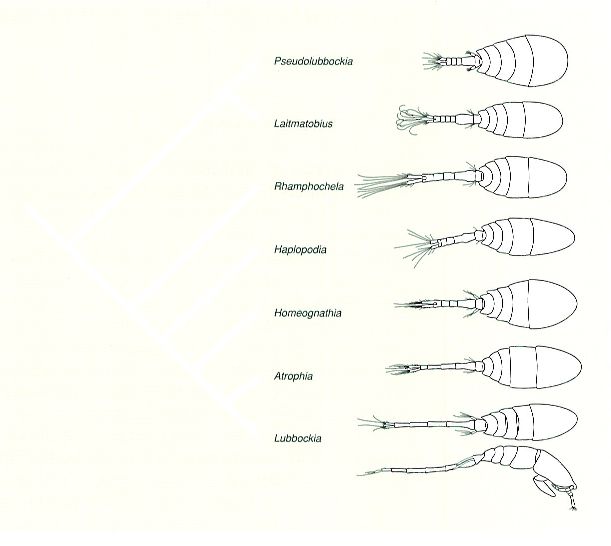
Cladograma mostrant les relacions entre els gèneres de lubòquids i la tendència evolutiva en la forma general del cos.
A partir de Huys i Böttger-Schnack, 1994/Barreras & Creixell.
L’any 1984 se celebrà a Ottawa la segona conferència internacional sobre copèpodes. Uns quants científics hi van exposar l’estat de la classificació dels diferents ordres de copèpodes amb vista a establir la filogènia del grup. S’arribà a la conclusió que l’anàlisi dels diferents caràcters corporals es pot emprar per a construir cladogrames que mostren com han evolucionat determinats caràcters bàsics. Segons quina sigui la variabilitat de cada caràcter (antenes, apèndixs toràcics o potes natatòries, etc.), es distingeix l’estat plesiomòrfic, o primitiu, de l’estat apomòrfic, o derivat. Per exemple, en els ciclopoides, el nombre de segments de l’antènula oscil·la entre 6 (en Cyclops) i 21 (en Euryte); la idea que preval en aquest cas és que com més elevat és el nombre de segments, més primitiu és l’estat. L’estat característic per als ciclopoides (Cyclopoida) és de 21 segments. Procedint d’aquesta manera es pot conèixer el caràcter més primitiu per a les antènules, les antenes, les mandíbules, les maxíl·lules, les maxil·les, els maxil·lípedes i els apèndixs toràcics o potes natatòries. Seguint aquest ordre d’idees, i segons els diversos cladogrames construïts, s’arriba a la conclusió que l’ordre filogenètic (de més primitiu a més modern), per exemple, seria el següent: ciclopínids (Cyclopinidae), ciclòpids (Cyclopoidae), oitònids (Oithonidae), arquinotododèlfids (Archinotododelphydae), notodèlfids (Notodelphydae), ascidiocòlids (Ascidiocolidae) i lerneids (Lernaeidae). També és interessant assenyalar l’aportació de les diverses tècniques moleculars a la filogènia del grup. Això ha fet que quan les darreres dècades s’han descrit noves espècies de vegades s’hagin estudiat utilitzant aquests mètodes cladístics (matrius de caràcters i cladogrames) i els caràcters plesiomòrfics o apomòrfics per a incloure-les en un dels grups. Això ha contribuït a separar diversos grups dins d’una mateixa família, i fins i tot a crear noves famílies, com és el cas dels onceids (Oncaeidae).
A continuació es presenta la classificació general dels copèpodes (a partir de Huys i Boxshall, 1991) i nombre d’espècies de cada ordre. Actualment s’estima que el nombre d’espècies sobrepassa les 11.200.
| Subclasse | Infraclasse | Superorders | Ordres | Medi | Hàbitat | Nombre d’espècies |
| Copèpodes (Copepoda) | Progimnopleus (Progimnoplea) | Progimnopleus (Progymnoplea) | Platicopioides (Platycopioida) | Marí | Planctònics hiperbentònics i d’aigües anquihalines | 11 |
| Neocopèpodes (Neocopepoda) | Gimnopleus (Gymnoplea) | Calanoides (Calanoida) | Marí i aigua dolça | Pelàgics i bentònics | 2.666 | |
| Podopleus (Podoplea) | Ciclopoides (Cyclopoida) | Marí i aigua dolça | Lliures, paràsits i comensals | 3.222 | ||
| Geliel·loides (Gelyelloida) | Aigües subterrànies | Sistemes càrstics | 3 | |||
| Harpacticoides (Harpacticoida) | Marí i aigua dolça | Bentònics, pelàgics i comensals | 3.643 | |||
| Misofrioides (Misophrioida) | Marí, anquihalí i aigua dolça | Aigües profundes i coves marines | 32 | |||
| Monstril·loides (Monstrilloida) | Marí | Nerítics | 74 | |||
| Mormonil·loides (Mormonilloida) | Marí | Pelàgics i paràsits | 2 | |||
| Pecilostomatoides (Poecilostomatoida) | Marí | Pelàgics | 254 | |||
| Sifonostomatoides (Siphonostomatoida) | Marí | Planctònics i paràsits | 1.641 |